Фосфатидилглицерол: Фосфатидилглицерол — Справочник химика 21
Фосфатидилглицерол — Справочник химика 21
Фосфатидилглицерол. Так же как фосфатидилинозитол, фосфотидил-глицерол не содержит азотсодержащего соединения. В этих фосфолипидах полярной группой служит еще одна молекула глицерола. [c.296]Фосфатидилглицеролы и их аминокислотные производные (например, ь-лизилфосфатидилглицерол) в значительном количестве содержатся в бактериальных мембранах, а также хлоропластах растений. [c.296]
Кардиолипин. Этот фосфолипид можно рассматривать как производное фосфатидилглицерола, в котором 3-гидроксигруппа второго остатка молекулы глицерола этерефицирована молекулой фосфатидной кислоты. [c.296]
ЦДФ-диацилглицерин можно рассматривать как переносчик фосфатидила. При переносе фосфатидила на спирт (инознтой, серин, глицерофосфат) образуются такие фосфоглицериды ЦДФ-диацилглицерин+инозитол фосфатидилинозит-[-ЦМФ ЦАФ-диацилглицерин серин фосфатидилсерин -f- ЦМФ 1ЩФ-диацилглице-рин — — глицерофосфат -> З-фосфатидилглицерол-1-фосфат -f- ЦМФ.
Активность пептидных антибиотиков определенным образом зависит от компонентного состава цитоплазматической мембраны и от трансмембранного электрического потенциала клетки-мишени. В составе бактериальных мембран присутствует большое количество кислых фосфолипидов (фосфатидилглицерола и кардиолипина), которые практически отсутствуют в мембранах эукариотических клеток. Эти фосфолипиды являются не только маркерами, но и центрами связывания пептидных антибиотиков животного происхождения, в частности дефенсинов и протегринов, за счет сильных ион-ионных связей, которые устанавливаются по кооперативному механизму аналогично концентрационной зависимости, представленной на рис. 10, А. Кроме того, известно, что мембранный потенциал бактерий в 1.5—2 раза выше, чем у мембран эукариотических клеток. Так что пептидные антибиотики с высоким содержанием лизина и аргинина могут проникать в клетку через мембрану путем электрофореза, подобно тому как это описано для микробных антибиотиков полимиксина В и грамицидинов (Франклин, Сноу, 1984).
В митохондриях присутствует фосфолипид кардиолипин (дифосфатидилглицерол). Он образуется из фосфатидилглицерола, который в свою очередь синтезируется из СОР-диацилглицерола (рис. 25.1) и глицерол-З-фосфата, как показано на схеме, приведенной на рис. 25.2.
Легочные сурфактанты » Справочник ЛС
Основное клиническое значение препаратов легочного сурфактанта определяется их эффективностью при тяжелых респираторных заболеваниях, связанных с первичным или вторичным дефицитом альвеолярного сурфактанта. К ним относятся следующие заболевания и синдромы:
Респираторный дистресс-синдром недоношенных новорожденных (РДСН), возникающий в результате незрелости легких и, как следствие, выраженного дефицита сурфактанта. Недостаточный синтез сурфактанта альвеолоцитами II типа и его быстрая инактивастного натяжения в альвеолах, их спадению (ателектазам), гиповентиляции и недостаточной оксигенации крови, развитию гиперкапнии и гипоксии.
Синдром аспирации мекония (САМ) у новорожденных возникает в результате попадания мекония в дыхательные пути, что приводит к нарушению синтеза сурфактанта, его инактивации компонентами мекония, спадению альвеол и развитию воспаления. Назначение сурфактанта способствует уменьшению тяжести заболевания, предупреждает развитие острой дыхательной недостаточности (ОДН).
Синдром острого повреждения легких (СОПЛ) и острый респираторный дистресс-синдром (ОРДС) взрослых — как наиболее тяжелая форма проявления СОПЛ. Легкие всегда вовлекаются в патологический процесс у больных, находящихся в критическом состоянии, вне зависимости от его этиологии. СОПЛ является основным компонентом полиорганной недостаточности, развивается в результате вторичного повреждения легких, включая все компоненты аэрогематического барьера и альвеолярного сурфактанта.
Пневмония сопровождается повреждением альвеолярной паренхимы, отеком легких с выходом белков плазмы крови в альвеолы, что оказывает угнетающее действие на активность сурфактанта и к спадению альвеол. Назначение сурфактанта в этих случаях приводит к улучшению газообмена, способствуя расправлению спавшихся альвеол, т.е. разрешению ателектазов.
Туберкулез легких, особенно его распространенные формы, вызывает глубокие нарушения в сурфактантной системе легких, обширные деструктивные и ателектатические изменения альвеолярной паренхимы, особенно в участках легкого, граничащих с крупными туберкулезными фокусами (инфильтраты, каверны и др.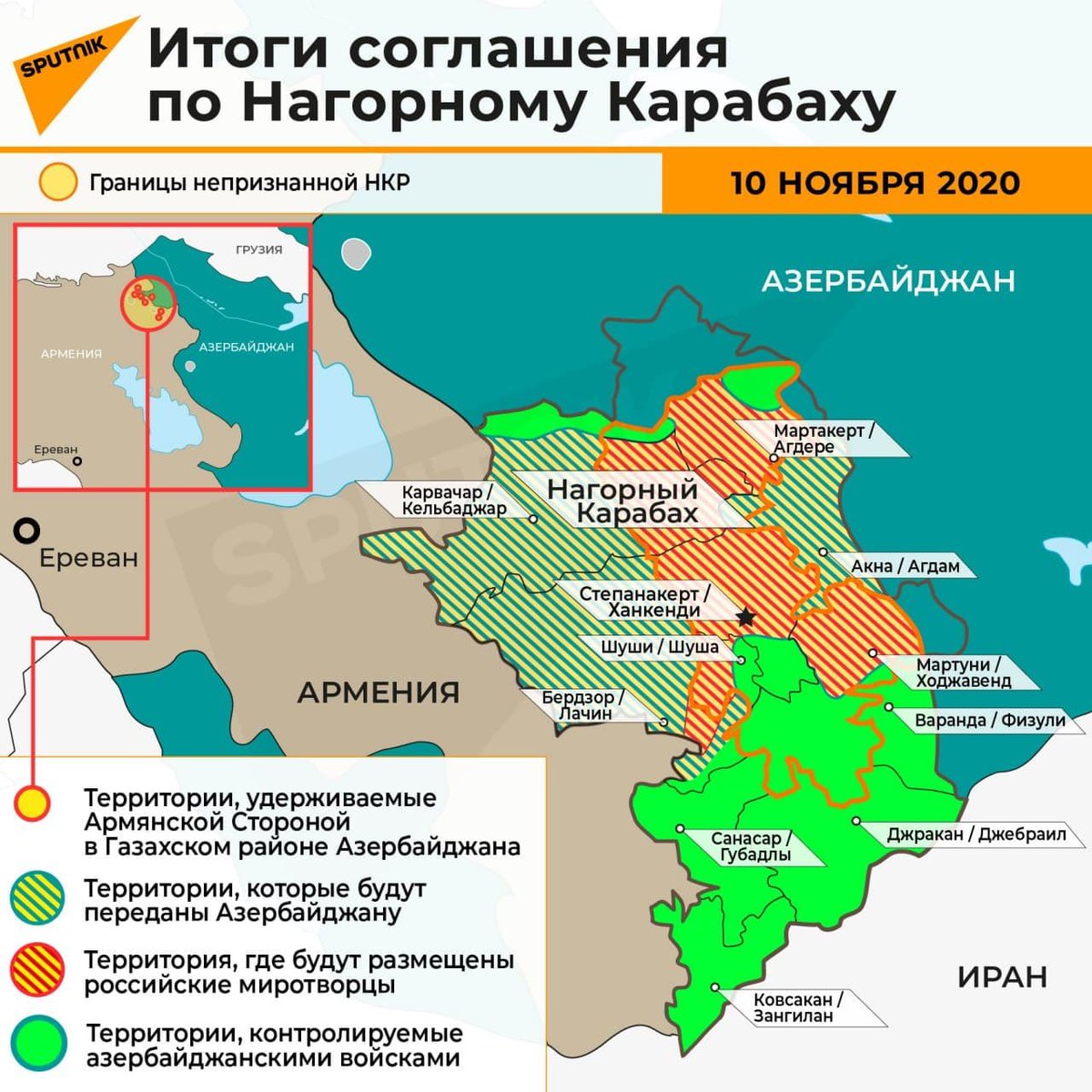 ). Применение препаратов сурфактанта в комплексном лечении туберкулеза позволяет уменьшить частоту возникновения сурфактант-зависимых изменений в легких, повысить поверхностно активные свойства легочного сурфактанта и способствует более быстрому обратному развитию воспалительных изменений.
). Применение препаратов сурфактанта в комплексном лечении туберкулеза позволяет уменьшить частоту возникновения сурфактант-зависимых изменений в легких, повысить поверхностно активные свойства легочного сурфактанта и способствует более быстрому обратному развитию воспалительных изменений.
В настоящее время разработаны основные принципы сурфактантотерапии:
- исчерпывающая базовая терапия;
- использование принципов неповреждающей механической вентиляции;
- раннее начало сурфактантотерапии (первые часы, сутки с момента развития РДС), при индексе оксигенации РаО2/ FiО2 ниже 200 мм рт. ст.;
- эндобронхиальное болюсное введение препарата в оптимальной дозе и повторное введение через 6—8 ч в случае положительного ответа на первое введение;
- сурфактантотерапия при положительном ответе на первое введение должна проводиться до достижения индекса оксигенации более 300 мм рт. ст., стойкого клинико-рентгенологического улучшения.
СИСТЕЙН Баланс ср-во офтал.
 10мл
10млСистейн® – система для комфорта глаз! Систейн® Баланс — это увлажняющие капли для глаз с системой Lipitech® (минеральное масло+анионы фосфолипидов), которая восстанавливает все слои слезной пленки. — устраняет слезотечение на ветер, солнце и мороз — устраняет «сухость глаз» при возрастных и гормональных изменениях — уменьшает кратность закапывания — увлажнение поверхности глаза; — стабилизация липидного слоя слезной пленки, уменьшение испарения слезной жидкости; — обеспечение временного облегчения признаков синдрома «сухого глаза» (чувство жжения, раздражение).
Индивидуальная повышенная чувствительность к компонентам препарата.
Применение при беременности и лактации
Безопасность и эффективность применения во время беременности и лактации не исследовались. Возможно применение при беременности и в период лактации по назначению лечащего врача, если ожидаемый эффект превышает риск развития возможных побочных эффектов.
Применение в педиатрии
Не рекомендуется применение в детском возрасте до 18 лет, так как безопасность и эффективность применения у детей не установлены.
Возможно применение при беременности и в период лактации по назначению лечащего врача, если ожидаемый эффект превышает риск развития возможных побочных эффектов.
Применение в педиатрии
Не рекомендуется применение в детском возрасте до 18 лет, так как безопасность и эффективность применения у детей не установлены.
Каждый миллилитр препарата содержит: пропиленгликоль; систему lipitech, включающую масло минеральное, димиристоил фосфатидилглицерол; гидроксипропилгуар; полиоксил стеарат; кислоту борную; сорбитантристеарат; сорбитол; поликвалид; динатрияздетат; хлористоводородную кислоту и/или гидроксид натрия; воду.
IMU49 — Антитела антифосфолипидные к кардиолипину / фосфатидной кислоте / фосфатидилхолину / фосфатидилэтаноламину / фосфатидилглицерину / фосфатидилинозитолу / фосфатидилсерину / аннексину V / β2-гликопротеину I / протромбину, IgM, раздельно
IMU49 — Антитела антифосфолипидные к кардиолипину / фосфатидной кислоте / фосфатидилхолину / фосфатидилэтаноламину / фосфатидилглицерину / фосфатидилинозитолу / фосфатидилсерину / аннексину V / β2-гликопротеину I / протромбину, IgM, раздельно — Заголовок сайта0
ВЫБРАННЫЕ ТЕСТЫ / УСЛУГИ
Антитела антифосфолипидные к кардиолипину / фосфатидной кислоте / фосфатидилхолину / фосфатидилэтаноламину / фосфатидилглицерину / фосфатидилинозитолу / фосфатидилсерину / аннексину V / β2-гликопротеину I / протромбину, IgM, раздельно
ИНТЕРПРЕТАЦИЯ РЕЗУЛЬТАТОВКардиолипин
♦ Основной лабораторный критерий АФС. Средняя и высокая концентрация антител указывает на наличие АФС. Высокий риск венозных тромбозов.
Средняя и высокая концентрация антител указывает на наличие АФС. Высокий риск венозных тромбозов.
Фосфатидная кислота, Фосфотидилхолин
♦ Ассоциированы с АФС. Часто встречаются в сочетании с антителами к кардиолипину и фосфатидилсерину
Фосфотидилэтаноламин
♦ Ассоциированы с АФС. Встречаются у 43% пациентов с АФС, в 5-17% отмечаются изолированно. Являются маркером серонегативного АФС.
Фосфатидилглицерол
♦ Ассоциированы с АФС, часто отмечаются вместе с другими отрицательно-заряженными фосфолипидами
Фосфотидилинозитол
♦ Встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. Являются доминантным маркером у молодых пациентов с церебральной ишемией, инсультом, мигренозными головными болями.
Фосфотидилсерин
♦ Являются маркером серонегативного АФС, коррелируют с волчаночным антикоагулянтом, реже встречаются при инфекционной патологии.
Аннексин V
♦ Являются некритериальным маркером АФС, часто отмечаются у больных с акушерской патологией на фоне аутоиммунных заболеваний. Тест целесообразен для анализа причины невынашивания.
β2-гликопротеин I
♦ Лабораторный критерий АФС. Средняя и высокая концентрация антител указывает на наличие АФС.
Протромбин
♦ Ассоциированы с клиническими проявлениями АФС и волчаночным антикоагулянтом.
- У детей до 16 лет забор биологического материала производится в присутствии родителей!
- За 1 день до сдачи анализа исключите физические нагрузки (спортивные тренировки) и прием алкоголя.
- Взятие крови предпочтительнее проводить утром натощак, после 8 — 14 часов ночного голодания (у маленьких детей – перед очередным грудным кормлением).
Сыворотка крови
Иммунодот, качественный тест
Медицинская лаборатория «Alfa Diagnostica». Все права защищены.
Все права защищены.
COVID-19 и ГРИППрегистрация ON-LINE
Тест на COVID-19от Мастеркард
keymeta
,keymeta
descmeta
descmeta
AIzaSyCT18YISEgdSRYzIM1Yuhq_OWkWdfrsCig
8189eea6a85da1b35815536ed6aca3ee
Антифосфолипидный синдром | Медицинский центр ЛМД
Антифосфолипидный синдром (АФС) — это аутоиммунное заболевание преимущественно женщин молодого возраста, характеризуемое рецидивирующими тромбозами, привычным невынашиванием беременности, тромбоцитопенией и стойким наличием антифосфолипидных антител. Диагноз АФС устанавливается на основании соответствия клинических и лабораторных находок диагностическим критериям этого заболевания.
Основу клинических проявлений АФС составляют повторные венозные тромбозы, локализуемые наиболее часто в глубоких венах голеней, что может осложняться тромбоэмболией легочной артерии. Однако тромбозы могут развиваться также в поверхностных венах и в любом другом венозном бассейне. Артериальные тромбозы проявляются гангреной конечностей, инсультами, окклюзией аорты с асептическим некрозом головок бедренной кости, инфарктом миокарда, почек и других внутренних органов. Тромбы могут образоваться на клапанах и в полостях сердца. Кожные проявления заключаются в язвах на голенях, кровоизлияниях в подногтевом ложе и сетчатым ливедо. Сетчатое ливедо в виде поверхностной синеватой венозной сеточки на голенях и бедрах особенно хорошо выявляется после охлаждения.Тромбоцитопения при АФС умеренная, редко приводит к осложнениям и не требует самостоятельной терапии. Идиопатическая тромбоцитопения, как и гемолитическая анемия является показанием к назначению лабораторных тестов, направленных на выявление антифосфолипидных антител. Необъяснимые спонтанные аборты на ранних сроках или внутриутробная гибель плода во 2-3 триметрах могут возникнуть при АФС без или на фоне тяжелой преэклампсии.
Артериальные тромбозы проявляются гангреной конечностей, инсультами, окклюзией аорты с асептическим некрозом головок бедренной кости, инфарктом миокарда, почек и других внутренних органов. Тромбы могут образоваться на клапанах и в полостях сердца. Кожные проявления заключаются в язвах на голенях, кровоизлияниях в подногтевом ложе и сетчатым ливедо. Сетчатое ливедо в виде поверхностной синеватой венозной сеточки на голенях и бедрах особенно хорошо выявляется после охлаждения.Тромбоцитопения при АФС умеренная, редко приводит к осложнениям и не требует самостоятельной терапии. Идиопатическая тромбоцитопения, как и гемолитическая анемия является показанием к назначению лабораторных тестов, направленных на выявление антифосфолипидных антител. Необъяснимые спонтанные аборты на ранних сроках или внутриутробная гибель плода во 2-3 триметрах могут возникнуть при АФС без или на фоне тяжелой преэклампсии.
Выделяют несколько форм АФС. Прежде всего, это первичный или идиопатический АФС, возникающий у лиц моложе 45 лет. После этого возраста большинство случаев рассматривается как вторичный АФС по отношению к основному заболеванию, обычно атеросклерозу и сердечной и венозной недостаточности. В то же время, обнаружение других факторов риска тромбоза не является поводом для исключения диагноза антифосфолипидного синдрома. Кроме того, у молодых лиц вторичный АФС развивается на фоне системной красной волчанки, синдрома Шегрена, склеродермии, опухолей, некоторых вирусных заболеваниях, лекарственной терапии.
Прежде всего, это первичный или идиопатический АФС, возникающий у лиц моложе 45 лет. После этого возраста большинство случаев рассматривается как вторичный АФС по отношению к основному заболеванию, обычно атеросклерозу и сердечной и венозной недостаточности. В то же время, обнаружение других факторов риска тромбоза не является поводом для исключения диагноза антифосфолипидного синдрома. Кроме того, у молодых лиц вторичный АФС развивается на фоне системной красной волчанки, синдрома Шегрена, склеродермии, опухолей, некоторых вирусных заболеваниях, лекарственной терапии.
В основе антифосфолипидного синдрома лежит появление антифосфолипидных антител (АФЛА). Они представляют собой гетерогенную популяцию иммуноглобулинов, взаимодействующих с отрицательно заряженными, реже с нейтральными фосфолипидами. АФС представляет собой иммуноопосредованое нарушение коагуляции, в ходе которого аутоантитела нарушают свертывание крови invitro, что в организме приводит к повторным тромбозам. Аутоантитела реагируют с фосфолипидами и их белковыми кофакторами, которые участвуют в образовании протромбинового комплекса каскада коагуляции. В результате удлиняется активированное частичное тромбопластиновое время (АЧТВ), которое отражает присутствие волчаночного антикоагулянта (ВАК).
Аутоантитела реагируют с фосфолипидами и их белковыми кофакторами, которые участвуют в образовании протромбинового комплекса каскада коагуляции. В результате удлиняется активированное частичное тромбопластиновое время (АЧТВ), которое отражает присутствие волчаночного антикоагулянта (ВАК).
Первые клинико-лабораторные критерии АФС были разработаны в 1998 году экспертами международного симпозиума в японском Саппоро. Они значительно ограничили клинические проявления, рассматриваемые как проявления АФС, а лабораторные критерии была основана на антителах к кардиолипину классов IgG и IgM и ВАК. В австралийском Сиднее в 2006 году произошел последний пересмотр критериев этого заболевания. Была несколько изменена трактовка клинических проявлений и в лабораторные критерии добавлены антитела к бета-2 гликопротеину I. На основе австралийских критериев в настоящее время строится практическая диагностика АФС.
Для иммунологического подтверждения АФС требуется в каждом случае проведение более одного теста, уровень антител должен был высоким, и эти значения должны сохраняться при повторном исследовании через 12 недель. Однако имеются исключения. Так, при привычном невынашивании беременности с прогностической целью достаточно определения антител к кардиолипину. Важно еще раз подчеркнуть, что согласно существующим критериям диагностики АФС для подтверждения этого аутоиммунного заболевания требуется обязательное сочетание хотя бы по одного иммунологического и одного клинического признака.
Однако имеются исключения. Так, при привычном невынашивании беременности с прогностической целью достаточно определения антител к кардиолипину. Важно еще раз подчеркнуть, что согласно существующим критериям диагностики АФС для подтверждения этого аутоиммунного заболевания требуется обязательное сочетание хотя бы по одного иммунологического и одного клинического признака.
Существует три общепринятых лабораторных теста, позволяющих выявить антитела, характерные для антифосфолипидного сиднома. К ним относят антитела к кардиолипину классов IgG и IgM, и антитела к бета-2 гликопротеину. Выявление ВАК является коагулогическим тестом. Назначение антикоагулянтов нередко препятствует обследованию пациентов в отношении ВАК.
Антитела к кардиолипину.
Кардиолипин (дифосфатидил глицерол) является отрицательно заряженным фосфолипидом обладающим значительной структурной гомологией с другими фосфолипидами (фосфатидилсерином, фосафтидилэтаноламином и другими), из которых состоят мембраны клеток человека. Кроме того, в комплексе с белками фосфолипиды принимают участие во многих рецепторных и ферментативных системах, в том числе системе свертывания крови. Антитела к кардиолипину относят к семейству антифосфолипидных антител.
Кроме того, в комплексе с белками фосфолипиды принимают участие во многих рецепторных и ферментативных системах, в том числе системе свертывания крови. Антитела к кардиолипину относят к семейству антифосфолипидных антител.
Наряду с антителами к бета-2-гликопротеину, антитела к кардиолипину входят в международные критерии антифосфолипидного синдрома 2006 года, а вместе с антителами к двуспиральной ДНК и Sm антигену в иммунологические критерии системной красной волчанки.
Мишенью антител к кардиолипину являются антигены, формирующиеся при объединении белковых ко-факторов и фосфолипидов. Основной мишенью антикардиолипиновых антител является неоантиген, образующийся при взаимодействии кардиолипина и сывороточного белка — бета-2-гликопротеина, который является естественным антикоагулянтом. Антитела к бета-2-гликопротеину отмечаются изолированно у 10-20% больных с АФС. В то же время, анти-кардиолипиновые антитела, направленные против фосфолипидов встречаются при сифилисе и ряде инфекционных и соматических заболеваний, поэтому малоспецифичны для диагноза антифосфолипидного синдрома.![]() Носительство антител к кардилипину при инфекциях имеет транзиторный характер, поэтому в таких случаях антитела отсутствуют при повторном определении через 3-6 месяцев.
Носительство антител к кардилипину при инфекциях имеет транзиторный характер, поэтому в таких случаях антитела отсутствуют при повторном определении через 3-6 месяцев.
В связи с вовлеченностью антител к кардиолипину во множество патогенетических процессов, их выявление имеет большое значение при АФС и ассоциированных состояниях. В патогенезе антифосфолипидного синдрома антитела in vitro нарушают образование протромбинового комплекса (протромбиназы), который состоит из фактора X, фактора V, фосфолипидов из тромбоцитов и кальция, что приводит к гипокоагуляции и удлинению времени основных коагуляционных тестов. Однако in vivo у больных АФС имеет место тромбообразование, генез которого окончательно не установлен. Кроме стимуляции протромботических механизмов, антикардиолипиновые антитела участвуют в индукции воспаления, нарушают функции эндотелия, нарушают функции трофобласта и плаценты, могут принимать участие в развитие атеросклероза.
Выявление антител к кардиолипину указывает на высокий риск развития системной красной волчанки. Антитела к кардиолипину классов IgG и IgM выявляются у 20-50% больных с системной красной волчанкой (СКВ) и 3-20% больных с другими системными ревматическими заболеваниями. СКВ является не единственным ревматологическим заболеванием, сопровождающимся синтезом антител к кардиолипидам. Сравнительно часто данные антитела находят у больных системной склеродермией, васкулитами и васкулитоподобными состояниями, например, при синдроме Бехчета и болезни Крона. Таким образом, клинические проявления антифосфолипидного синдрома на фоне другого ревматического заболевания указывают на «вторичный» антифосфолипидный синдром. Для его выявления целесообразно дообследовать пациентов для выявления антинуклеарного фактора на НЕр2 клеточной линии. Отдельную клиническую форму АФС представляет собой фульминантный «катастрофический» АФС, характеризующийся множественными тромбозами и быстрым развитием ДВС синдрома. У всех больных с катастрофическим антифосфолипидным синдромом отмечаются крайне высокие титры антител к кардиолипину.
Антитела к кардиолипину классов IgG и IgM выявляются у 20-50% больных с системной красной волчанкой (СКВ) и 3-20% больных с другими системными ревматическими заболеваниями. СКВ является не единственным ревматологическим заболеванием, сопровождающимся синтезом антител к кардиолипидам. Сравнительно часто данные антитела находят у больных системной склеродермией, васкулитами и васкулитоподобными состояниями, например, при синдроме Бехчета и болезни Крона. Таким образом, клинические проявления антифосфолипидного синдрома на фоне другого ревматического заболевания указывают на «вторичный» антифосфолипидный синдром. Для его выявления целесообразно дообследовать пациентов для выявления антинуклеарного фактора на НЕр2 клеточной линии. Отдельную клиническую форму АФС представляет собой фульминантный «катастрофический» АФС, характеризующийся множественными тромбозами и быстрым развитием ДВС синдрома. У всех больных с катастрофическим антифосфолипидным синдромом отмечаются крайне высокие титры антител к кардиолипину. При отсутствии системного ревматического заболевания антифосфолипидный синдром рассматривают как первичный АФС.
При отсутствии системного ревматического заболевания антифосфолипидный синдром рассматривают как первичный АФС.
Антитела к бета-2-гликопротеин I.
Бета-2-гликопротеин I представляет собой гликозилированный белок плазмы крови, содержание которого в норме составляет около 150-300 мг/л. Хотя его функции окончательно не установлены, однако известно, что он обладает выраженной антикоагулянтной активностью. Немаловажной функцией бета-2-гликопротеина является его способность связываться с апоптотическими тельцами и участвовать в их клиренсе. Именно участие бета-2-гликопротеина в процессах апоптоза может быть одним из основных механизмов, приводящих к сенсибилизации иммунной системы аутоантигеном, что приводит к таким аутоиммунным заболеваниям как АФС и системная красная волчанка. В 2006 году определение антител к бета-2-гликопротину-1 было включено в лабораторные классификационые критерии антифосфолипидного синдрома.
Бета-2-гликопротеин I является белковым кофактором антител к кардиолипину и образует неоантиген, который распознается бета2-гликопротеин зависимыми антителами к кардиолипину. Этот антигенный комплекс является основной серологической мишенью при АФС. В эксперименте иммунизация бета-2-гликопротеином индуцирует характерную картину с появлением антител к кардиолипину, значительному увеличению числа резорбций плода, тромбоцитопении и удлинению времени ряда коагулогических тестов.
Этот антигенный комплекс является основной серологической мишенью при АФС. В эксперименте иммунизация бета-2-гликопротеином индуцирует характерную картину с появлением антител к кардиолипину, значительному увеличению числа резорбций плода, тромбоцитопении и удлинению времени ряда коагулогических тестов.
Связывание бета-2-гликопротеину I на пластике иммуноферментного планшета приводит к экспозиции скрытых эпитопов молекулы. Антитела к бета-2-гликопротеину I являются единственным серологическим маркером АФС у значимой части пациентов. Диагностическая важность выявления антител к бета-2-гликопротеину I в патогенезе АФС подтвержадается его включением в число лабораторных классификационных критериев АФС.
В тест системах для выявления антител к кардиолипину первого поколения использовался бета2 гликопротеин сыворотки крупного рогатого скота, в то время как в тест-системах второго поколения используется рекомбинантный человеческий белок.
Антитела к фосфатидилсерин-протромбиновому комплексу.
Антитела к фосфатидилсерин-протромбиновому комплексу относятся к семейству антифосфолипидных антител. Фосфатилилсерин представляет собой основной отрицательно заряженный фосфолипид клеточных мембран. Кроме того, фосфатидилсерин является кофактором бета-2-гликопротеина I , протромбина, тромбомодулина и ряда других фосфолипид-зазависимых белков принимающих участие в каскаде свертывания.
Протромбин (фактор II) представляет собой витамин К-зависимый гликопротеин с концентрацией в плазме 100 мг/л. Трансформация протромбина в тромбин происходит под действием протромбинового комплекса, состоящего из факторов коагуляции Xa, Vaи ионов Ca2+.
Фосфатидилсериновый-протромбиновый (PS-PT) комплекс является одной из мишеней антифосфолипидных антител. Аутоантитела к фосфатидилсерин-протромбиновому комплексу играют роль в нарушении каскада коагуляции, принимают большое развитии артериальных и венозных тромбозов у больных с антифосфолипидным синдромом.
Антитела к фосфатидилсерин-протромбиновому комплексу отмечаются у 40-60% больных с классическим антифосфолипидным синдромом, кроме того могут встречаться при «серонегативном» некритериальном антифосфолипидном синдроме. Антитела отмечаются при высоком риске артериального тромбоза, кроме того их выявление может быть одним из факторов риска сосудистых тромбозов при системных ревматических заболеваниях.
Выявление этой разновидности антител хорошо коррелирует с результатами выявления ВАК, поэтому выявление антител к фосфатидилсерин-протромбиновому комплексу может подтверждать наличие ВАК в случае назначения антикоагуляционной терапии.
Антитела к аннексину V.
Аннексин V представляет собой кальций-зависимый белок, связывающийся с анионными фосфолипидами (фосфатидилсерином, кардиолипином ). Он связан на мембранах клеток, которые находятся в контакте с плазмой крови, в том числе эндотелиоцитах, тромбоцитах и клетках трофобласта. Благодаря его способности образовать внутреннюю «выстилку» сосудов, на поверхности эндотелиоцитов аннексин V проявляет выраженную антикоагулянтную активность. За счет формирования защитного антикоагулянтный слоя на поверхности эндотелиоцитов и ворсин трофобласта, обращенных в просвет сосуда, поддерживается целостность эндотелиальных мембран.
При АФС аннексин V является одним из кофакторов фосфолипидных антител. Антитела к фосфолипидам способны связываться с аннексином, что приводит к разрушению защитной выстилки, в результате чего значительно повышается риск тромбозов и нарушается микроциркуляция. Особенно чувствительны к антителам к аннексину V клетки трофобласта. Антитела приводят к нарушению формирования плаценты, снижают синтез ряда гормонов и факторов синцитиотрофобластом.
Кроме того, показана способность антител к аннексину V принимать участие в нарушениях микроциркуляции при ряде аутоиммунных состояний, включая повторные выкидыши при АФС и системной красной волчанке, поражения сосудов при аортоартериите Такаясу, легочной гипертензии при склеродермии.
Клинические исследования указывают, что клинико-диагностические параметры тестов по обнаружению антител к аннексину V не позволяют их использовать для оценки риска невынашивания в общей популяции, однако тест целесообразен для анализа причины невынашивания у больных с аутоиммунными заболеваниями (например, больных СКВ).
Иммуноблот антифосфолипидных антител.
К антифосфолипидным антителами относятся антититела к негативно-заряженным фосфолипидам — кардиолипину, фосфатидилхолину, фосфатидилэтаноламину, фосфатидилглицеролу, фосфатидилэназитолу, фосфатидилсерину, антитела к их кофакторам — бета-2-гликопротеину I, аннексину V и протромбину и функциональный коагулогический тест — ВАК. Патогенные свойства антифосфолипидных антител обусловлены их протромбогенным потенциалом, индукцией воспаления, нарушением функции эндотелия, функции трофобласта и плаценты, участием в развитие атеросклероза. Так как антифосфолипидные антитела встречаются при других аутоиммунных, инфекционных и лимфопролиферативных заболеваний, их выявление малоспецифично для диагноза АФС. Носительство антител при инфекциях имеет транзиторный характер, поэтому в таких случаях антитела отсутствуют при повторном определении через 3-6 месяцев.
Диагностика АФС, основанная только на одном из трех лабораторных критериев, может быть ошибочна. Это утверждение подтверждается мета-анализом, показывающим, что присутствие только антител к кардиолипину и бета-2-гликопротеину I само по себе не связано с тромбоэмболическими событиями. При этом важно исследовать спектр антифосфолипидных антител, позволяющий установить позитивность по нескольким маркерам, которая более точно выявляет риск повторных тромбозов и тромбоэмболий, а также патологию беременности.
Основным отличием иммуноблота антифосфолипидных антител от твердофазных ИФА тест-систем является пористая структура гидрофобной PVDF-мембраны, которая ориентирует гидрофильные участки фосфолипидов и обеспечивает тем самым более плотное их распределение. Мембранная иммобилизация такого рода имитирует представление антигена на анионных фосфолипидах in vivo и может улучшить связывание антител.
Также тест является комплексным и позволяет детектировать не только к кардиолипину и бета-2-гликопротеину I, но и антитела к фосфатидилхолину, фосфатидилэтаноламину, фосфатидилглицеролу, фосфатидилиназитолу, фосфатидилсерину, аннексину V и протромбину IgG и IgM.
Антиген аутоантител и класс антител | Значение |
Кардиолипин (дифосфатидил-глицерол) ,антитела класса IgG | Основной лабораторный показатель при АФС. Содержание антител в среднем и высоком титре указывает на наличие АФС. |
Фосфатидная кислота,антитела класса IgG | Ассоциированы с АФС. Часто встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. |
Фосфатидилхолин ,антитела класса IgG | Ассоциированы с АФС. Часто встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. |
Фосфатидилэтаноламин, антитела класса IgG | Ассоциированы с АФС. Встречаются у 43% пациентов с АФС, в 5-17% отмечаются изолированно. Являются маркером серонегативного АФС. |
Фосфатидилглицерол, антитела класса IgG | Ассоциированы с АФС, часто отмечаются вместе с другими отрицательно-заряженными фосфолипидами |
Фосфатидилинозитол, антитела класса IgG | Встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. Являются доминантным маркером у молодых пациентов с церебральной ишемией, инсультом, мигренозными головными болями. |
Фосфатидилсерин, антитела класса IgG | Являются маркером серонегативного АФС, коррелируют с волчаночным антикоагулянтом, реже встречаются при инфекционной патологии. |
Аннексин V, антитела класса IgG | Являются некритериальным маркером АФС, часто отмечаются у больных с акушерской патологией на фоне аутоиммунных заболеваний. Тест целесообразен для анализа причины невынашивания. |
Бета-2 гликопротеин 1, антитела класса IgG | Лабораторный критерий АФС. Содержание антител в среднем и высоком титре указывает на наличие АФС. Антитела класса IgGотмечаются у больных с высоким риском артериальных тромбозов. |
Протромбин, антитела класса IgG | Ассоциированы с клиническими проявлениями АФС и волчаночным антикоагулянтом. Отмечаются у больных с высоким риском тромбозов. |
Кардиолипин (дифосфатидил-глицерол), антитела класса IgM | Лабораторный критерий АФС. Содержание антител в среднем и высоком титре указывает на наличие АФС. Антитела класса IgM отмечаются у пациентов с риском венозных тромбозов. |
Фосфатидная кислота, антитела класса IgM | Ассоциированы с АФС. Часто встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. |
Фосфатидилхолин, антитела класса IgM | Ассоциированы с АФС. Часто встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. |
Фосфатидилэтаноламин, антитела класса IgM | Ассоциированы с АФС. Встречаются у 43% пациентов с АФС, в 5-17% — изолированно. Являются маркером серонегативного АФС. |
Фосфатидилглицерол, антитела класса IgM | Ассоциированы с АФС. |
Фосфатидилинозитол, антитела класса IgM | Встречаются в сочетании с антителами к кардилипину и фосфатидилсерину. Являются доминантным маркером у молодых пациентов с церебральной ишемией |
Фосфатидилсерин, антитела класса IgM | Являются маркером серонегативного АФС, коррелируют с волчаночным антикоагулянтом, реже встречаются при инфекционной патологии. |
Аннексин V, антитела класса IgM | Являются некритериальным маркером АФС. Тест целесообразен для анализа причины невынашивания у больных с аутоиммунными заболеваниями. |
Бета-2 гликопротеин 1, антитела класса IgM | Лабораторный критерий АФС. Содержание антител в среднем и высоком титре указывает на наличие АФС. |
Протромбин, антитела класса IgM | Ассоциированы с клиническими проявлениями АФС и волчаночным антикоагулянтом. |
Документы
Биология для студентов — 09. Фосфо
Фосфолипиды. Общий признак всех фосфолипидов — наличие в их составе фосфорной кислоты. В зависимости от спиртового компонента они делятся на глицерофосфолипиды и сфингофосфолипиды.
Глицерофосфолипиды. Общим структурным фрагментом всех глицерофосфолипидов является фосфатидная кислота (1,2-диацил,3-фосфоглицерол):
Молекулы фосфолипидов имеют гидрофобную часть, образованную радикалами жирных кислот, и гидрофильную — остатки фосфорной кислоты, аминокислот, аминоспиртов.
Фосфатидилхолин (лецитин). В своем составе содержит аминоспирт холин (гидроксид 3-гидроксиэтилтриметиламмония). В зависимости от того, с каким атомом углерода глицерола связана фосфорная кислота (a— в крайнем положении, b — в срединном), различают два типа фосфатидилхолинов (a и b).
Фосфатидилхолины (ФХ) широко распространены в клетках, особенно мозговой ткани человека и животных; в растениях они встречаются в соевых бобах, семенах подсолнечника, зародышах пшеницы.
Фосфатидилэтаноламин (кефалин). В состав фосфатидилэтаноламинов (ФЭА) вместо холина входит азотистое основание этаноламин.
ФХ и ФЭА являются главными липидными компонентами мембран клеток.
В результате гидролиза при действии специфичного фермента — фосфолипазы А2 в ФХ или ФЭА возможно отщепление остатка жирной кислоты с образованием лизоформ: лизофосфатидилхолина (ЛФХ) и лизофосфатидилэтаноламина (ЛФЭА):
Фосфатидилсерин. В молекуле фосфатидилсерина полярной группой является остаток аминокислоты серина.
Фосфатидилсерины (ФС) распространены менее широко, чем ФХ и ФЭА. Значение ФС определяется тем, что он является предшественником в синтезе двух других групп.
Плазмалогены. Известны также глицерофосфолипиды, которые в отличие от приведенных выше — ФХ, ФЭА, ФС — отличаются тем, что вместо остатка кислоты при атоме углерода С, они содержат a-, b-ненасыщенный спирт, образующий простую эфирную связь с гидроксильной группой глицерола.
При гидролизе этой эфирной связи образуется альдегид соответствующего спирта. Отсюда название группы — фосфатидали: фосфатидальэтаноламин, фосфатидальхолин, фосфатидальсерин.
На долю плазмалогенов приходится до 10% фосфолипидов мозга и мышечной ткани. Они обнаружены также в эритроцитах, тканях некоторых беспозвоночных (до 25%), входят в состав бактериальных мембран, но практически не встречаются в растениях.
Фосфатидилинозитол. В отличие от других групп глицерофосфолипидов в состав фосфатидилинозитола вместо азотсодержащих соединений входит шестиуглеродный циклический спирт инозитол, представленный одним из его стереоизомеров — моноинозитолом.
Фосфатидилинозитол входит в состав клеточных мембран животных, высших растений, различных микроорганизмов, особенно высоко его содержание в миелиновых оболочках нервных волокон.
Важное значение имеют фосфорилированные производные фосфатидилинозитолов.
Фосфатидилглицерол. Так же как фосфатидилинозитол, фосфотидилглицерол не содержит азотсодержащего соединения. В этих фосфолипидах полярной группой служит еще одна молекула глицерола.
Фосфатидилглицеролы и их аминокислотные производные в значительном количестве содержатся в бактериальных мембранах, а также хлоропластах растений.
Кардиолипин. Этот фосфолипид можно рассматривать как производное фосфатидилглицерола, в котором 3-гидроксигруппа второго остатка молекулы глицерола этерефицирована молекулой фосфатидной кислоты.
Кардиолипин локализован почти исключительно в митохондриях и играет важную роль в структурной организации и функционировании дыхательных комплексов. Кардиолипин является также обязательным компонентом бактериальных клеточных мембран и хлоропластов растений.
Существенная роль фосфатидилглицерина в фотосинтезе
Andersson B, Aro E-M (2001) Фотоповреждения и оборот белка D1 в фотосистеме II. В: Aro E-M, Andersson B (eds) Регулирование фотосинтеза. Kluwer Academic Publishers, Дордрехт, Нидерланды, стр. 377–393
Google Scholar
Эндрюс Дж., Мадд Дж. Б. (1985) Синтез фосфатидилглицерина в хлоропластах гороха. Путь и локализация. Физиология растений 79: 259–265
PubMed CAS Google Scholar
Aro E-M, Virgin I, Andersson B (1993) Фотоингибирование фотосистемы II.Инактивация, повреждение белков и обмен. Biochim Biophys Acta 1143: 113–134
PubMed Статья CAS Google Scholar
Бабийчук Э., Мюллер Ф., Эубель Х., Браун Х-П, Френцен М., Кушнир С. (2003) Arabidopsis фосфатидилглицерофосфатсинтаза 1 важна для дифференцировки хлоропластов, но необязательна для митохондриальной функции. Завод J 33: 899–909
PubMed Статья CAS Google Scholar
Бенсон А.А., Маруо Б. (1958) Фосфолипиды растений.Идентификация фосфатидилглицеринов. Biochim Biophys Acta 27: 189–195
PubMed Статья CAS Google Scholar
Бенсон А.А., Маруо Б. (1989) «Нова» в метаболизме фосфатов, GPG и открытие фосфатидилглицерина. Biochim Biophys Acta 1000: 447–451
PubMed CAS Google Scholar
Блок М.А., Дорн А.Дж., Джоярд Дж., Дус Р. (1983) Получение и характеристика мембранных фракций, обогащенных внешней и внутренней оболочкой мембран из хлоропластов шпината: II.Биохимическая характеристика. J Biol Chem 258: 13281–13286
PubMed CAS Google Scholar
Browse J, McCourt P, Somerville CR (1985) Мутант Arabidopsis , лишенный хлоропласт-специфичного липида. Science 227: 763–765
Статья CAS PubMed Google Scholar
Browse J, Somerville C (1991) Синтез глицеролипидов — биохимия и регуляция.Annu Rev Plant Physiol Plant Mol Biol 42: 467–506
Артикул CAS Google Scholar
Карман Г.М., Генри С.А. (1999) Биосинтез фосфолипидов в дрожжах Saccharomyces cerevisiae и взаимосвязь с другими метаболическими процессами. Prog Lipid Res 38: 361–399
PubMed Статья CAS Google Scholar
Demonkos I, Malec P, Sallai A, Kovács L, Itoh K, Shen G, Ughy B, Bogos B, Sakurai I, Kis M, Strzalka K, Wada H, Itoh S, Farkas T, Gombos Z (2004 г. ) Фосфатидилглицерин необходим для олигомеризации реакционного центра фотосистемы I.Plant Physiol 134: 1471–1478
Статья CAS Google Scholar
Дорн А.Дж., Джойард Дж., Дус Р. (1990) Действительно ли тилакоиды содержат фосфатидилхолин? Proc Natl Acad Sci USA 87: 71–74
PubMed Статья CAS Google Scholar
Dowhan W (1997) Молекулярная основа разнообразия мембранных фосфолипидов: почему так много липидов? Анну Рев Биохим 66: 199–232
PubMed Статья CAS Google Scholar
Droppa M, Horváth G, Hideg E, Farkas T (1995) Роль фосфолипидов в регуляции фотосинтетической активности транспорта электронов: обработка тилакоидов фосфолипазой C.Photosynth Res 46: 287–293
Статья CAS Google Scholar
Dubertret G, Mirshahi A, Mirshahi M, Gerard-Hirne C, Tremolieres A (1994) Данные in vivo манипуляций с липидным составом у мутантов, что Δ 3 — транс, содержащий -гексадеценовую кислоту фосфатидилглицерин участвует в биогенезе светособирающего хлорофилла a / b -белкового комплекса Chlamydomonas reinhardtii .Eur J Biochem 226: 473–482
PubMed Статья CAS Google Scholar
Dubertret G, Gerard-Hirne C, Trémolières A (2002) Важность trans -Δ 3 -гексадеценовая кислота, содержащая фосфатидилглицерин в образовании тримерного светособирающего комплекса в Chlamydomonas . Plant Physiol Biochem 40: 829–836
Статья CAS Google Scholar
Frentzen M (2004) Фосфатидилглицерин и сульфохиновозилдиацилглицерин: анионные мембранные липиды и регуляция фосфатов.Curr Opin Plant Biol 7: 270–276
PubMed Статья CAS Google Scholar
Frentzen M, Heinz E, McKeon TA, Stumpf PK (1983) Специфичность и селективность глицерин-3-фосфатацилтрансферазы из хлоропластов гороха и шпината. Eur J Biochem 129: 629–636
PubMed CAS Статья Google Scholar
Frentzen M, Nishida I, Murata N (1987) Свойства пластидного ацил- (белка-носителя ацила): глицерин-3-фосфатацилтрансферазы из чувствительного к холоду растения кабачка ( Cucurbita moschata ).Физиология растительных клеток 28: 1195–1201
CAS Google Scholar
Gombos Z, Várkonyi Z, Hagio M, Iwaki M, Kovács L, Masamoto K, Itoh S, Wada H (2002) Требование фосфатидилглицерина для функции акцептора электронов пластохинона Q B в реакционном центре фотосистемы II. Биохимия 41: 3796–3802
PubMed Статья CAS Google Scholar
Грибау Р., Френцен М. (1994) Биосинтез фосфатидилглицерина в изолированных митохондриях этиолированных проростков маша ( Vigna radiata L).Физиология растений 105: 1269–1274
PubMed CAS Google Scholar
Hagio M, Gombos Z, Várkonyi Z, Masamoto K, Sato N, Tsuzuki M, Wada H (2000) Прямые доказательства потребности фосфатидилглицерина в фотосистеме II фотосинтеза. Физиол растений 124: 795–804
PubMed Статья CAS Google Scholar
Hagio M, Sakurai I, Sato S, Kato T, Tabata S, Wada H (2002) Фосфатидилглицерин необходим для развития тилакоидных мембран у Arabidopsis thaliana .Физиология растительных клеток 43: 1456–1464
PubMed Статья CAS Google Scholar
Hobe S, Prytulla S, Kühlbrandt W, Paulsen H (1994) Тримеризация и кристаллизация восстановленного светособирающего комплекса хлорофилла a / b . EMBO J 13: 3423–3429
PubMed CAS Google Scholar
Hobe S, Förster R, Klingler J, Paulsen H (1995) Мотив N-проксимальной последовательности в светособирающем хлорофилле a / b -связывающем белке важен для тримеризации светособирающего хлорофилла a / b комплекс.Биохимия 34: 10224–10228
PubMed Статья CAS Google Scholar
Ishizaki O, Nishida I, Agata K, Eguchi G, Murata N (1988) Клонирование и нуклеотидная последовательность кДНК пластидной глицерин-3-фосфатацилтрансферазы из тыквы. FEBS Lett 238: 424–430
PubMed Статья CAS Google Scholar
Ishizaki-Nishizawa O, Fujii T, Azuma M, Sekiguchi K, Murata N, Ohtani T., Toguri T (1996) Низкотемпературная устойчивость высших растений значительно усиливается неспецифической цианобактериальной десатуразой.Nat Biotechnol 14: 1003–1006
PubMed Статья CAS Google Scholar
Джордан Б.Р., Чоу В.С., Бейкер А.Дж. (1983) Роль фосфолипидов в молекулярной организации мембран хлоропластов гороха: влияние истощения фосфолипидов на фотосинтетическую активность. Biochim Biophys Acta 725: 77–86
Статья CAS Google Scholar
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauß N (2001) Трехмерная структура фотосистемы I цианобактерий в точке 2.Разрешение 5 Å. Nature 411: 909–917
PubMed Статья CAS Google Scholar
Joyard J, Maréchal E, Miege C, Block MA, Dorne A-J, Douce R (1998) Структура, распределение и биосинтез глицеролипидов из хлоропластов высших растений. В: Siegenthaler P-A, Murata N (eds) Липиды в фотосинтезе. Kluwer Academic Publishers, Дордрехт, Нидерланды, стр. 21–52
Google Scholar
Kenrick JR, Bishop DG (1986) Фосфатидилглицерин и сульфохиновозилдиацилглицерин в листьях и плодах чувствительных к холоду растений.Фитохимия 25: 1293–1295
Статья CAS Google Scholar
Kruse O, Schmid GH (1995) Роль фосфатидилглицерина как функционального эффектора и мембранного якоря пептида ядра D1 из частиц фотосистемы II цианобактерии Oscillatoria chalybea . Z Naturforsch 50c: 380–390
Google Scholar
Kruse O, Hankamer B, Konczak C, Gerle C, Morris E, Radunz A, Schmid GH, Barber J (2000) Фосфатидилглицерин участвует в димеризации фотосистемы II.J Biol Chem 275: 6509–6514
PubMed Статья CAS Google Scholar
Kurisu G, Zhang H, Smith JL, Cramer WA (2003) Структура цитохрома b 6 f Комплекс кислородного фотосинтеза: настройка полости. Science 302: 1009–1014
PubMed Статья CAS Google Scholar
Liu Z, Yan H, Wang K, Kuang T, Zhang J, Gui L, An X, Chang W. (2004) Кристаллическая структура основного светособирающего комплекса шпината на 2.Разрешение 72 Å. Nature 428: 287–292
PubMed Статья CAS Google Scholar
Loll B, Kern J, Seanger W, Zouni A, Biesiadka J (2005) К полному расположению кофакторов в структуре с разрешением 3,0 Å фотосистемы II. Nature 438: 1040–1044
PubMed Статья CAS Google Scholar
Lyons JK (1973) Переохлаждение растений. Annu Rev Plant Physiol 24: 445–466
Артикул CAS Google Scholar
Maanni AE, Dubertret G, Delrieu MJ, Roche O, Trémolières A (1998) Мутанты Chlamydomonas reinhardtii , затронутые метаболизмом фосфатидилглицерина и биогенезом тилакоидов.Plant Physiol Biochem 36: 609–619
Статья Google Scholar
Малкин Р., Нийоги К. (2000) Фотосинтез. В: Buchanan BB, Gruissem W, Jones RL (eds) Биохимия и молекулярная биология растений. Американское общество физиологов растений, Роквилл, Мэриленд, стр. 568–628
Google Scholar
Мацумото К. (2001) Обязательная природа фосфатидилглицерина в Escherichia coli : двойная роль анионных фосфолипидов.Mol Microbiol 39: 1427–1433
PubMed Статья CAS Google Scholar
McCourt P, Browse J, Watson J, Arntzen CJ, Somerville CR (1985) Анализ функции фотосинтетической антенны у мутанта Arabidopsis thaliana (L.), лишенного транс -гексадеценовой кислоты. Физиология растений 78: 853–858
PubMed CAS Google Scholar
Мелис А. (1991) Динамика состава и функции фотосинтетических мембран.Biochim Biophys Acta 1058: 87–106
Статья CAS Google Scholar
Moon BY, Higashi S, Gombos Z, Murata N (1995) Ненасыщенность мембранных липидов хлоропластов стабилизирует фотосинтетический аппарат от низкотемпературного фотоингибирования у трансгенных растений табака. Proc Natl Acad Sci USA 92: 6219–6223
PubMed Статья CAS Google Scholar
Мур Т.С. мл. (1974) Синтез фосфатидилглицерина в эндосперме клещевины.Кинетика, требования и внутриклеточная локализация. Физиология растений 54: 164–168
PubMed CAS Google Scholar
Мур Т.С. мл. (1982) Биосинтез фосфолипидов. Annu Rev Plant Physiol 33: 235–259
Артикул CAS Google Scholar
Mudd JB, Dezacks R (1981) Синтез фосфатидилглицерина хлоропластами из листьев Spinacia oleracea L.(шпинат). Arch Biochem Biophys 209: 584–591
PubMed Статья CAS Google Scholar
Murata N (1983) Молекулярный видовой состав фосфатидилглицеринов из чувствительных к холоду и устойчивых к холоду растений. Физиология растительных клеток 24: 81–86
CAS Google Scholar
Murata N, Nishida I (1987) Липиды сине-зеленых водорослей (цианобактерии). В: Штумпф П.К., Конн Е.Е. (ред.) Биохимия растений, том 9.Academic Press, Орландо, США, стр. 315–347
Google Scholar
Murata N, Tasaka Y (1997) Глицерин-3-фосфат ацилтрансфераза в растениях. Biochim Biophys Acta 1348: 10–16
PubMed CAS Google Scholar
Мурата Н., Вада Х. (1995) Ацил-липидные десатуразы и их значение в устойчивости и акклиматизации цианобактерий к холоду. Biochem J 308: 1–8
PubMed CAS Google Scholar
Мурата Н., Ямая Дж. (1984) Температурно-зависимое фазовое поведение фосфатидилглицеринов из чувствительных к холоду и устойчивых к холоду растений.Физиология растений 74: 1016–1024
PubMed CAS Статья Google Scholar
Мурата Н., Сато Н., Такахаши Н., Хамазаки Ю. (1982) Состав и позиционное распределение жирных кислот в фосфолипидах из листьев чувствительных к холоду и устойчивых к холоду растений. Физиология растительных клеток 23: 1071–1079
CAS Google Scholar
Murata N, Wada H, Gombos Z (1992a) Режимы десатурации жирных кислот у цианобактерий.Физиология растительных клеток 33: 933–941
CAS Google Scholar
Мурата Н., Ишизаки-Нисидзава О, Хигаши С., Хаяси Х, Тасака И., Нисида I (1992b) Генетически модифицированное изменение чувствительности растений к холоду. Nature 356: 710–713
Статья CAS Google Scholar
Нишида И., Мурата Н. (1996) Чувствительность к холоду растений и цианобактерий: решающий вклад липидов мембран.Annu Rev Plant Physiol Plant Mol Biol 47: 541–568
PubMed Статья CAS Google Scholar
Nishida I, Frentzen M, Ishizaki O, Murata N (1987) Очистка изомерных форм ацил- (белка-носителя ацила): глицерин-3-фосфатацилтрансферазы из зеленеющих семядолей кабачков. Физиология растительных клеток 28: 1071–1079
CAS Google Scholar
Nishida I, Tasaka Y, Shiraishi H, Murata N (1993) Ген и РНК предшественника глицерин-3-фосфатацилтрансферазы, расположенной в пластиде Arabidopsis Талиана .Plant Mol Biol 21: 267–277
PubMed Статья CAS Google Scholar
Нишияма Ю., Аллахвердиев С.И., Мурата Н. (2006) Новая парадигма действия активных форм кислорода при фотоингибировании фотосистемы II. Biochim Biophys Acta 1757: 742–749
PubMed Статья CAS Google Scholar
Nußberger S, Dörr K, Wang DN, Kühlbrandt W (1993) Липидно-белковые взаимодействия в кристаллах светособирающего комплекса растений.J Mol Biol 234: 347–356
PubMed Статья Google Scholar
Ohlrogge J, Browse J (1995) Биосинтез липидов. Растительная ячейка 7: 957–970
PubMed Статья CAS Google Scholar
Ohnishi M, Thompson Jr GA (1991) Биосинтез уникального компонента trans -Δ 3 -гексадеценовой кислоты фосфатидилглицерина хлоропластов: данные о его месте и механизме образования.Arch Biochem Biophys 288: 591–599
PubMed Статья CAS Google Scholar
Okazaki K, Sato N, Tsuji N, Tsuzuki M, Nishida I (2006) Значение жирных кислот C 16 в положениях sn -2 глицеролипидов в фотосинтетическом росте Synechocystis sp. PCC6803. Физиология растений 141: 546–556
PubMed Статья CAS Google Scholar
Филлипс М.С., Хаузер Х., Палтауф Ф. (1972) Меж- и внутримолекулярное смешение углеводородных цепей в системах лецитин / вода.Chem Phys Lipids 8: 127–133
PubMed Статья CAS Google Scholar
Raison JK (1973) Влияние фазовых изменений, вызванных температурой, на кинетику дыхательных и других мембраносвязанных ферментов. J Bioenerg 4: 285–309
PubMed Статья CAS Google Scholar
Raison JK, Wright LC (1983) Термические фазовые переходы в полярных липидах мембран растений.Их индукция диненасыщенными фосфолипидами и их возможная связь с переохлаждением. Biochim Biophys Acta 731: 69–78
Статья CAS Google Scholar
Roughan G, Slack R (1984) Синтез глицеролипидов в листьях. Trends Biochem Sci 9: 383–386
Статья CAS Google Scholar
Roughan PG, Thompson GA Jr, Cho SH (1987) Метаболизм экзогенных длинноцепочечных жирных кислот листьями шпината.Arch Biochem Biophys 259: 481–496
PubMed Статья CAS Google Scholar
Sakamoto A, Sulpice R, Kaneseki T, Hou C-X, Kinoshita M, Higashi S, Moon BY, Nonaka H, Murata N (2004) Генетическая модификация ненасыщенности жирных кислот хлоропластного фосфатидилглицерина изменяет чувствительность к холодовому стрессу. Среда растительной клетки 27: 99–105
Статья CAS Google Scholar
Sakurai I, Hagio M, Gombos Z, Tyystjärvi T, Paakkarinen V, Aro E-M, Wada H (2003) Требование фосфатидилглицерина для обслуживания фотосинтетического оборудования.Физиология растений 133: 1376–1384
PubMed Статья CAS Google Scholar
Сакураи И., Шен Дж.Р., Ленг Дж., Охаши С., Кобаяши М., Вада Х. (2006) Липиды в кислородно-выделяющих комплексах фотосистемы II цианобактерий и высших растений. J Biochem 140: 201–209
PubMed Статья CAS Google Scholar
Sato N (2004) Роль кислых липидов сульфохиновозилдиацилглицерина и фосфатидилглицерина в фотосинтезе: их специфичность и эволюция.J Plant Res 117: 495–505
PubMed Статья CAS Google Scholar
Сато Н., Мурата Н. (1982a) Биосинтез липидов у сине-зеленой водоросли Anabaena variabilis . I. Липидные классы. Biochim Biophys Acta 710: 271–278
CAS Google Scholar
Сато Н., Мурата Н. (1982b) Биосинтез липидов в сине-зеленых водорослях, Anabaena variabilis .II. Жирные кислоты и молекулы липидов. Biochim Biophys Acta 710: 279–289
CAS Google Scholar
Сато Н., Хагио М., Вада Х, Тузуки М. (2000) Потребность в фосфатидилглицерине для фотосинтетической функции в тилакоидных мембранах. Proc Natl Acad Sci USA 97: 10655–10660
PubMed Статья CAS Google Scholar
Сато Н., Суда К., Цузуки М. (2004) Ответственность фосфатидилглицерина за биогенез комплекса PSI.Biochim Biophys Acta 1658: 235–243
PubMed Статья CAS Google Scholar
Schlame M, Rua D, Greenberg ML (2000) Биосинтез и функциональная роль кардиолипина. Prog Lipid Res 39: 257–288
PubMed Статья CAS Google Scholar
Shibuya I (1992) Регуляция метаболизма и биологические функции фосфолипидов в Escherichia coli .Prog Lipid Res 31: 245–299
PubMed Статья CAS Google Scholar
Siegenthaler P-A (1998) Молекулярная организация ациллипидов в фотосинтетических мембранах высших растений. В: Siegenthaler P-A, Murata N (eds) Липиды в фотосинтезе. Kluwer Academic Publishers, Дордрехт, Нидерланды, стр. 119–144
Google Scholar
Somerville C, Browse J, Jaworski JG, Ohlrogge JB (2000) Липиды.В: Buchanan BB, Gruissem W, Jones RL (eds) Биохимия и молекулярная биология растений. Американское общество физиологов растений, Мэриленд, стр. 456–527
Google Scholar
Stroebel D, Choquest Y, Popot JL, Picot D (2003) Атипичный гем в цитохроме b 6 ф комплекс. Nature 426: 413–418
PubMed Статья CAS Google Scholar
Szalontai B, Kota Z, Nonaka H, Murata N (2003) Структурные последствия генетически модифицированного насыщения жирных кислот фосфатидилглицерина в тилакоидных мембранах табака.Исследование FTIR. Биохимия 42: 4292–4299
PubMed Статья CAS Google Scholar
Tasaka Y, Nishida I, Higashi S, Beppu T, Murata N (1990) Жирнокислотный состав фосфатидилглицеринов в зависимости от чувствительности древесных растений к холоду. Физиология растительных клеток 31: 545–550
CAS Google Scholar
Trémolières A, Siegenthaler P-A (1998) Восстановление липидами.В: Siegenthaler P-A, Murata N (eds) Липиды в фотосинтезе. Kluwer Academic Publishers, Дордрехт, Нидерланды, стр. 175–189
Google Scholar
Wada H, Murata N (1989) Synechocystis Мутанты PCC6803, дефектные по десатурации жирных кислот. Физиология растительных клеток 30: 971–978
CAS Google Scholar
Wada H, Murata N (1998) Мембранные липиды у цианобактерий.В: Siegenthaler P-A, Murata N (eds) Липиды в фотосинтезе. Kluwer Academic Publishers, Дордрехт, Нидерланды, стр. 65–81
Google Scholar
Weier D, Müller C, Gaspers C, Frentzen M (2005) Характеристика ацилтрансфераз из Synechocystis sp. PCC6803. Biochem Biophys Res Commun 334: 1127–1134
PubMed Статья CAS Google Scholar
Wolter FP, Schmidt R, Heinz E (1992) Чувствительность к холоду Arabidopsis thaliana с генетически модифицированными мембранными липидами.EMBO J 11: 4685–4692
PubMed CAS Google Scholar
Xu C, Härtel H, Wada H, Hagio M, Yu B, Eakin C, Benning C (2002) Мутантный локус pgp1 Arabidopsis кодирует фосфатидилглицерофосфатсинтазу с нарушенной активностью. Физиология растений 129: 594–604
PubMed Статья CAS Google Scholar
Yu B, Xu C, Benning C (2002) Arabidopsis , разрушенный в SQD2. , кодирующий сульфолипидсинтазу, нарушается при ограниченном фосфатом росте.Proc Natl Acad Sci USA 99: 5732–5737
PubMed Статья CAS Google Scholar
Cellsome® — на основе фосфатидилглицерина (PG) — Encapsula NanoSciences
Описание
Простые липосомы широко используются в качестве моделей искусственных клеток и для изучения взаимодействия различных типов молекул, таких как пептиды и белки, с поверхностью липидных мембран. Простые липосомы также используются в качестве контрольных составов для многих различных типов липосомальных составов с инкапсулированным лекарственным средством.Заряженные липосомы также используются во многих исследованиях комплемента крови. Простые липосомы доступны с различными анионными, катионными и нейтральными поверхностными зарядами. Простые липосомы могут быть однослойными или многослойными.
Фосфатидилсерин (PS) и фосфатидилглицерин (PG) — две хорошо известные анионные молекулы, которые образуют липосомы с отрицательным дзета-потенциалом. Дзета-потенциал липосом варьируется в зависимости от молярного процента отрицательно заряженных липидов в липосомальной композиции.Каталоги Anionic Cellsome®-Phosphatidylserine (PS) и Cellsome®-Phosphatidylglycerol (PG) содержат продукты, содержащие от 0,5 до 100 процентов DOPS и DOPG, соответственно, с различным количеством отрицательных дзета-потенциалов.
Строение молекулы ДОПГ с анионной головной группой.Загрузить вкладыш продукта Загрузить паспорт безопасности (SDS)
Информация о составе
Cellsome® — на основе фосфатидилглицерина (PG)
Для получения дополнительной информации о липидном составе упомянутых выше липосом щелкните здесь.
| Размер буфера и липосомы | Спецификация |
|---|---|
| Буфер | Физиологический раствор с фосфатным буфером * |
| pH | 7,4 |
| Размер липосом | 100 нм |
| * Если вы предпочитаете другие буферы, укажите это при заказе. | |
Калькулятор частиц липосом
Липосомына основе Cellsome®-фосфатидилглицерина (PG) являются однослойными и имеют размер до 100 нм.Молярная концентрация липосом составляет 10 мМ. Имея диаметр липосом (нм) и концентрацию липидов (мкМ), вы можете рассчитать общее количество липидов в одной липосоме и количество липосом в одном миллилитре раствора липосом. Чтобы воспользоваться калькулятором, нажмите здесь.
Каждая липосома размером 100 нм состоит из почти 80 000 молекул фосфолипидов. В 10 мМ растворе липосом (концентрация фосфолипидов 10 мМ) содержится 75,2 триллиона (75,2 × 10 12 ) липосомных частиц на миллилитр раствора.При использовании калькулятора липосом необходимо помнить о двух вещах:
- Введите концентрацию в мкМ. Например, 10 мМ раствор составляет 10 000 мкМ.
- Включите в расчет только концентрацию фосфолипидов. Холестерин не учитывается, поскольку он встраивается в цепи гидрофобных жирных кислот липида.
Этот расчет работает только для однослойных липосом размером менее 200 нм. Он не работает с более крупными многослойными липосомами.Подробнее о расчете смотрите в видео ниже:
Технические примечания
- DOPG и DOPC являются ненасыщенными липидами, и температура их фазового перехода из жидкости в гель ниже нуля градусов Цельсия.
- DOPG и DOPC имеют хвосты ненасыщенных жирных кислот. У ненасыщенного липида есть «извилистые ножки», поэтому он образует неплотно упакованные липосомы. Насыщенные липиды образуют плотно упакованные липосомы. Упаковка липосом будет влиять на взаимодействие молекул с поверхностью липосом.
- Липосомы являются однослойными и имеют размер до 100 нм. Если вам нужно, чтобы они были другого размера, сообщите об этом при заказе. Однослойные липосомы могут быть изготовлены с размерами 50 нм и 200 нм. Липосомы размером более 200 нм имеют многослойную структуру.
- Липосомы изготавливаются в буфере PBS при pH 7,4, но их можно приготовить в любом другом буфере по вашему выбору. Вы можете указать свой буфер при заказе.
- Липосомы изготавливаются в дегазированном буфере, который продувают аргоном, чтобы избежать окисления ненасыщенных фосфолипидов.
- Липосомы производятся в стерильных условиях.
- Липосомы являются однослойными, поэтому из-за своего небольшого размера они не оседают на дне флакона.
- Если вам нужно взять несколько аликвот из флакона для использования в течение определенного периода времени, рекомендуется проявлять особую осторожность, чтобы не загрязнить флакон. Рекомендуется хранить флакон под стерильным колпаком для сохранения стерильности продукта.
- Липосомы нельзя замораживать.Кристаллы льда, образующиеся при замораживании, разрывают липидную мембрану липосом и изменяют размер липосомных частиц.
- Липосомы следует хранить в холодильнике при 4 ° C, чтобы избежать гидролиза липосом.
Внешний вид
Cellsome® — на основе фосфатидилглицерина (PG) — это белая полупрозрачная жидкость, состоящая из однослойных липосом наноразмеров. Обычно из-за небольшого размера липосом на дне флакона не происходит оседания. Липосомы расфасованы в ампулы янтарного цвета.
Информация для заказа / доставки
- Все составы на основе липосом отправляются на голубом льду при 4 ° C в изолированных упаковках с использованием ночной доставки или международной экспресс-доставки.
- Липосомы НИКОГДА нельзя замораживать. Кристаллы льда, образующиеся в липидной мембране, могут разорвать мембрану и изменить размер липосом. Липосомы в жидком виде всегда следует хранить в холодильнике.
- Клиенты, которые делают заказы из-за пределов Соединенных Штатов Америки, несут ответственность за свои государственные налоги на импорт и таможенные документы.Encapsula NanoSciences НЕ несет ответственности за импортные сборы в страны за пределами Соединенных Штатов Америки.
- Мы настоятельно рекомендуем клиентам в Японии, Корее, Тайване и Китае делать заказы через дистрибьюторов. Жесткие правила таможенного оформления в этих странах приведут к задержкам в таможенном оформлении этих скоропортящихся продуктов, если они заказываются напрямую через нас. Дистрибьюторы могут легко очистить посылки на таможне. Чтобы увидеть список дистрибьюторов, нажмите здесь.
- Клиенты, заказывающие у университетов и исследовательских институтов в Австралии, должны помнить, что липосомальные препараты сделаны из синтетического материала, и для составов не требуется «разрешение на импорт карантинного материала».Липосомы НЕ являются биологическими продуктами.
- Если вы хотите, чтобы со счета FedEx или DHL вашего учреждения взималась плата за доставку, укажите номер счета при оформлении заказа.
- Encapsula NanoSciences не контролирует задержки из-за плохой погоды или задержек при таможенном оформлении. После подтверждения заказа вы получите номер для отслеживания FedEx или DHL. Свяжитесь с FedEx или DHL заранее и убедитесь, что оформление документов для таможни оформлено вовремя. Все последующие запросы о доставке следует направлять в Federal Express или DHL.
Хранение и срок годности
Хранилище
ПродуктыCellsome® следует всегда хранить в темноте при 4 ° C, за исключением случаев, когда они на короткое время доводятся до комнатной температуры перед дозированием животным. НЕ МЕРЗНИ. Если суспензия заморожена, инкапсулированное лекарство может высвобождаться из липосом, что ограничивает его эффективность. Кроме того, размер липосом также изменится при замораживании и оттаивании.
Срок годности
Cellsome® — на основе фосфатидилглицерина (PG) производится ежедневно.Отгруженная партия изготавливается в тот же день. Рекомендуется использовать продукцию в течение 4 месяцев с даты изготовления.
Ссылки и справочная информация
1. Папагеоргиу Ф., Пиппа Н., Назирис Н., Деметзос С. Физико-химическое исследование белок-липосомных взаимодействий: влияние состава и концентрации липосом на связывание с белками. Журнал липосомных исследований. 23 апреля 2018 г. (только что принято): 1-29 .
2. Chonn A, Semple SC, Cullis PR.Ассоциация белков крови с большими однослойными липосомами in vivo. Отношение к срокам обращения. Журнал биологической химии. 1992 15 сентября; 267 (26): 18759-65 .
3. Semple SC, Chonn A, Cullis PR. Взаимодействие липосом и систем-носителей на основе липидов с белками крови: связь с поведением клиренса in vivo. Расширенные обзоры доставки лекарств. 1998 июн 8; 32 (1-2): 3-17 .
4. Девайн Д.В., Вонг К., Серрано К., Чонн А., Каллис ПР.Взаимодействие липосом и комплемента в сыворотке крови крыс: значение для исследований выживаемости липосом. Biochimica et Biophysica Acta (BBA) -Biomembranes. 1994 20 апреля; 1191 (1): 43-51 .
5. Szebeni J. Взаимодействие липосом с системой комплемента. Critical Reviews ™ в терапевтических системах-носителях лекарственных средств. 1998; 15 (1) .
6. Себени Дж., Бараньи Л., Савай С., Милошевиц Дж., Бодо М., Бангер Р., Алвинг ЧР. Взаимодействие липосом с системой комплемента: анализы in vitro и in vivo.InMethods в энзимологии, 1 января 2003 г. (том 373, стр. 136-154). Академик Пресс .
7. Бонте Ф., Джулиано Р.Л. Взаимодействие липосом с белками сыворотки крови. Химия и физика липидов. 1 июня 1986 г .; 40 (2-4): 359-72 .
8. Chonn A, Semple SC, Cullis PR. Ассоциация белков крови с большими однослойными липосомами in vivo. Отношение к срокам обращения. Журнал биологической химии. 1992 15 сентября; 267 (26): 18759-65 .
9. Колтье Дж. П., Шейз Б., Винтерхальтер М., Фурнье Д. Инкапсуляция белка в липосомах: эффективность зависит от взаимодействий между белком и бислоем фосфолипидов. Биотехнология BMC. 2002 декабрь; 2 (1): 9 .
10. Грегориадис Г., редактор. Липосомная технология: взаимодействие липосом с биологической средой. CRC пресс; 2006 12 сентября .
11. Papahadjopoulos D, Kimelberg HK. Фосфолипидные везикулы (липосомы) как модели биологических мембран: их свойства и взаимодействие с холестерином и белками.Прогресс в науке о поверхности. 1974, 1 января; 4: 141-232 .
12. Kimelberg HK. Взаимодействие белков и липосом и их значение для структуры и функции клеточных мембран. Молекулярная и клеточная биохимия. 1 февраля 1976 г .; 10 (3): 171-90 .
Потребность в фосфатидилглицерине для фотосинтетической функции тилакоидных мембран
Реферат
Для исследования роли фосфатидилглицерина (PG) в фотосинтезе мы сконструировали мутант, дефектный по гену CDP-диацилглицеринсинтазы, из цианобактерии Synechocystis sp.PCC6803. Мутант, обозначенный как SNC1, требовал добавления PG для роста. Рост подавлялся в среде без PG одновременно с уменьшением клеточного содержания PG. Эти результаты показывают, что PG важен, и что SNC1 нарушает синтез PG. Снижение содержания PG сопровождалось уменьшением клеточного содержания хлорофилла, но с небольшим влиянием на содержание пигментов фикобилисом, что показало, что уровни хлорофилл-белковых комплексов снижались без изменения уровней фикобилисом.Несмотря на снижение содержания PG, CO 2 -зависимый фотосинтез SNC1 был подобен фотосинтезу дикого типа на основе хлорофилла, но, следовательно, стал ниже на клеточной основе. Одновременно, отношение выделения кислорода фотосистемы II (ФСII), измеренное с p -бензохиноном, к таковому для CO 2 -зависимого фотосинтеза, которое колебалось между 1,3 и 1,7 у дикого типа. Однако он был снижен в SNC1 с 1,3 до 0,4 во время ранней фазы роста, когда содержание хлорофилла и CO 2 -зависимый фотосинтез были мало затронуты, а затем, наконец, до 0.1, предполагая, что ФСII сначала потеряла способность восстанавливать p -бензохинон, а затем снизилась в своем уровне и фактической активности. Эти результаты показывают, что PG вносит вклад в накопление хлорофилл-белковых комплексов в тилакоидных мембранах, а также в нормальное функционирование ФСII.
Биологические мембраны со специальными функциями демонстрируют специфический белковый и липидный состав. Мембранные липиды образуют барьер между внутренней и внешней стороной мембран, снабжают мембранные белки местами для их действия и регулируют функции мембранных белков.До сих пор исследования функционирования мембранных систем были сосредоточены в основном на самих белках, обладающих фактической каталитической активностью, и в гораздо меньшей степени на липидах мембран, которые окружают или интегрированы в мембранные белки. Исследования последнего типа включают ( i ) изучение эффектов удаления или добавления определенных липидов на некоторые очищенные белковые комплексы мембран, ( ii ) анализ липидов, связанных с комплексами, и ( iii ) определение характеристик мембранных функций мутантов, дефектных по определенному липиду.Хотя первые две стратегии привели к накоплению значительного количества информации о возможной роли липидов, окончательный вывод об их роли in vivo не может быть сделан из-за потенциальных артефактов, возникающих, например, при использовании липидов. моющие средства с экспериментальных систем in vitro . Напротив, последняя стратегия была успешно использована для получения in vivo доказательств роли липидов.
Тилакоидные мембраны, сайты для фотосинтетического транспорта электронов и фотофосфорилирования, содержат моногалактозилдиацилглицерин (MGDG), дигалактозилдиацилглицерин (DGDG), сульфохиновозилдиацилглицерин (SQDG), являющийся основным компонентом элевации.г., 1). Мутанты, лишенные SQDG и DGDG, были выделены и охарактеризованы для некоторых фотосинтезирующих организмов. Мутант, лишенный SQDG зеленой водоросли, Chlamydomonas reinhardtii , показал снижение активности фотосистемы II (PSII) на ≈40% с небольшим повреждением активности фотосистемы I (PSI), несмотря на незначительное влияние на содержание комплекса PSII или его субъединичный состав (2, 3). Вместе с наблюдением специфической ассоциации SQDG с комплексом PSII, но не с комплексом PSI, у C.reinhardtii , результаты можно интерпретировать как демонстрацию вклада SQDG в полную экспрессию активности PSII посредством поддержания нормальной конформации комплекса PSII посредством специфического связывания (3). С другой стороны, мутант высшего растения Arabidopsis thaliana с 90% снижением содержания ДГДГ проявлял комплексные эффекты на фотосинтез (т. Е. Увеличение размера антенны ФСII и уменьшение соотношения ФСII / ФСI ), что предполагает участие ДГДГ в структурной организации фотосинтетического аппарата (4, 5).
Для выяснения функции PG, единственного фосфолипида в мембранах тилакоидов, необходимо получить мутанты, дефектные по PG. Однако определение физиологической роли PG посредством анализа мутантов до сих пор было успешным только для нефотосинтезирующих организмов (6-14). Например, нулевая мутация фосфатидилглицеринфосфат (PGP) синтазы привела к потере синтезов PG и кардиолипина (CL) и, в конечном итоге, к летальному исходу у Escherichia coli и Saccharomyces cerevisiae в условиях неферментационного роста (9, 14).Эти наблюдения можно объяснить заселением большей части анионных липидов PG и / или CL в мембранных системах E. coli и митохондриях эукариотических клеток (14, 15). Недавно сообщалось, что нуль-мутант E. coli стал жизнеспособным на фоне дефицита основного липопротеина внешних мембран (16). Анионные липидные составляющие фотосинтетических мембранных систем отличаются от таковых нефотосинтетических мембран E.coli и S. cerevisiae , состоящие из PG и нефосфолипида (SQDG). Таким образом, представляет особый интерес установить, является ли PG незаменимым для фотосинтезирующих организмов. CDP-диацилглицеринсинтаза (CDS), локализованная в пути биосинтеза PG, была охарактеризована для нескольких организмов посредством клонирования генов и / или кДНК. Поиск сходства ДНК-последовательности генома цианобактерии Synechocystis sp. PCC6803 выявил одну ORF, кодирующую гомолог CDS (17).Мы получили мутант с дефектом синтеза PG из Synechocystis sp. PCC6803 посредством разрушения этой ORF посредством гомологичной рекомбинации и охарактеризовали эффекты этой мутации на фотосинтетический аппарат.
Материалы и методы
Клеточная культура.
Клетки дикого типа и мутантные клетки Synechocystis sp. PCC6803 выращивали фотоавтотрофно в среде BG-11 (18) при постоянном освещении люминесцентной лампой (10 Вт⋅м -2 ) при 30 ° C, в продолговатых стеклянных сосудах, аэрированных воздухом, содержащим 2% CO 2 , или в колбах. с тряской.Диолеоил-PG (Sigma) обрабатывали ультразвуком в среде BG-11 для получения липосом (6 мМ) для фильтрации для стерилизации. В культуральную среду добавляли липосомы PG в указанных концентрациях, где указано.
Выделение ДНК.
Клетки Synechocystis sp. PCC6803 выращивали, как описано выше, и собирали центрифугированием (8000 × g, , 10 мин, 4 ° C). ДНК выделяли из этих клеток, как описано в ссылке. 19.
Амплификация предполагаемого гена CDS
Synechocystis sp.PCC6803.Для амплификации кодирующей области предполагаемого гена CDS, обнаруженного в геноме Synechocystis sp. PCC6803, ПЦР выполняли с праймером 1 (5′-GGGTCATATGCCCACCCAGCGAATTATTAG-3 ‘) и праймером 2 (5′-GGGTGGATCCTCACAAATTATTCAACACAG-3’). Подчеркнутые кодонов в праймерах 1 и 2 являются стартовыми и стоп-кодонами соответственно. ПЦР выполняли с помощью термоциклера ДНК (Perkin-Elmer) со следующей программой термоцикла: 2 мин при 95 ° C, затем 25 циклов по 30 с при 95 ° C, 60 с при 55 ° C и 90 с при 72 ° C. ° C.Основной продукт размером 1,1 т.п.н. очищали из агарозного геля после электрофореза.
Скрининг библиотеки геномной ДНК
Synechocystis sp. PCC6803.Библиотека геномной ДНК Synechocystis sp. PCC6803 был сконструирован в бактериофаге λDASHII (Stratagene) путем интеграции геномной ДНК, частично расщепленной с помощью Sau 3AI, в фаговый вектор. Геномную ДНК подвергали скринингу с использованием предполагаемого гена CDS в качестве зонда посредством гибридизации бляшек, как описано ранее (20).
Конструирование мутанта с разрушением предполагаемого гена CDS.
Плазмиду, содержащую предполагаемый разрушенный ген CDS, использовали для трансформации дикого типа Synechocystis sp. PCC6803 путем гомологичной рекомбинации, как описано в ссылке. 21. Разрушение предполагаемого гена подтверждали с помощью ПЦР с праймерами 1 и 2, описанными выше.
Анализ липидов.
Общие липиды экстрагировали из клеток и затем разделяли на отдельные классы липидов с помощью двумерной ТСХ, как описано ранее (2).Пятна липидов визуализировали опрыскиванием примулином. Затем получали метиловые эфиры жирных кислот из общих липидов и каждого класса липидов с 5% безводной HCl в метаноле для анализа с помощью капиллярной ГЖХ, как описано ранее (2). Содержание липидов оценивали с использованием арахидоновой кислоты в качестве внутреннего стандарта.
Определение содержания пигментов и фотосинтеза.
Содержание хлорофилла определяли после его экстракции из клеток 100% метанолом (22), тогда как содержание фикоцианина, аллофикоцианина и фикоэритрина определяли, как описано в ссылке.23. Фотосинтетический O 2 эволюция в сочетании с CO 2 фиксацию измеряли в среде BG-11, содержащей клетки, эквивалентные 2,67 мкг Chl⋅ml -1 и 10 мМ NaHCO 3 с электродом типа Кларка Rank Brothers, Кембридж, Великобритания). Электродную камеру заполняли 5 мл реакционной смеси, выдерживали при 30 ° C и освещали вольфрамовой лампой проектора (180 Вт 180м -2 ).
Результаты и обсуждение
Нарушение предполагаемого гена CDS в
Synechocystis sp.PCC6803.Посредством поиска сходства последовательности ДНК генома цианобактерии Synechocystis sp. PCC6803, одна ORF, slr1369, как было обнаружено, кодирует предполагаемую CDS, состоящую из 293 аминокислот с молекулярной массой 31,9 кДа (16). Аминокислотная последовательность, выведенная из этой открытой рамки считывания, проявляла 38% идентичность с последовательностью CDS E. coli и 27-30% идентичностью с последовательностями CDS S. cerevisiae , Drosophila , крыса и A.thaliana (ссылки 24–28 и данные не показаны). Предполагаемые CDS Synechocystis sp. PCC6803, как и CDS E. coli и эукариотических организмов, обладал последовательностью, соответствующей мотиву CDS, SMMKRDAGVKDSGQLIPGHGGILDRTD, на своем С-конце (база данных PROSITE, http://www.expasy.ch/sprot/prosite.html ), и несколько потенциальных областей, перекрывающих мембрану (данные не показаны), как обнаружено при анализе гидрофобности в соответствии с методом Кайта и Дулиттла (29). С другой стороны, гидрофильный N-конец, обычно присутствующий в эукариотических CDS, не обнаружен ни в последовательности цианобактерий, ни в последовательности E.coli CDS (данные не показаны). Эти особенности предполагают, что данная ORF действительно кодирует CDS Synechocystis sp. PCC6803.
Затем мы сконструировали нулевой мутант предполагаемого гена CDS Synechocystis sp. PCC6803 для определения роли этого предполагаемого гена в отношении липидных и фотосинтетических свойств. Фрагмент ДНК размером 0,9 т.п.н., соответствующий slr1369, амплифицировали с помощью ПЦР с геномной ДНК Synechocystis sp. PCC6803 в качестве матрицы, а затем лигировали с pUC119 с получением плазмиды pCDSP.Вставка pCDSP показала сайты рестрикции, ожидаемые от последовательности генома Synechocystis sp. PCC6803 (данные не показаны), и его использовали в качестве зонда для скрининга библиотеки геномной ДНК Synechocystis sp. PCC6803 построен в λDASH II. В результате мы получили клон фага λ, содержащий вставку размером 14 т.п.н. Ожидалось, что фрагмент Bam HI / Hin dIII размером 4,4 т.п.н. во вставке будет содержать предполагаемый ген CDS из геномной последовательности Synechocystis sp.PCC6803. Затем мы вырезали фрагмент размером 4,4 т.п.н. для лигирования в pUC119 с получением плазмиды pCDS1. Было подтверждено, что вставка pCDS1 содержит предполагаемый ген с его 5′- и 3′-фланкирующими областями с помощью рестрикционного картирования (данные не показаны). Затем pCDS1 разрезали с помощью Bsm I / Bam HI, полученный фрагмент размером 2,4 т.п.н. затупляли и снова лигировали в pUC119 с получением плазмиды pCDS2 (фиг. 1 a ). Плазмида pCDS2 была расщеплена Bal I для удаления фрагмента 0,5 т.п.н. в предполагаемом гене CDS, который был заменен на 1.Фрагмент размером 4 т.п.н., несущий кассету устойчивости к канамицину (Km r ) pHSG298 для продукции плазмиды pCDS3 (фиг. 1 a ). Нарушение предполагаемого гена дикого типа Synechocystis sp. PCC6803 был проведен с pCDS3 посредством гомологичной рекомбинации в присутствии липосом PG в случае потребности в PG мутантом для его роста, что было проверено с помощью ПЦР с праймерами для 5′- и 3′-концевых областей ORF (рис. 1 б ). А 0.Фрагмент размером 9 т.п.н. амплифицировали, как ожидалось для дикого типа, тогда как с мутантом наблюдали только фрагмент размером 1,8 т.п.н., соответствующий гену ORF∷Km r , что доказывает полную замену ORF на ORF∷Km . г ген. Мы обозначили нулевой мутант предполагаемого гена CDS как SNC1.
Рисунок 1Конструирование разрушителя slr1369 из Synechocystis sp. PCC6803. ( a ) Конструирование плазмиды для разрушения slr1369 в Synechocystis sp.PCC6803. ( b ) Подтверждение с помощью ПЦР разрушения slr1369 путем вставки гена устойчивости к канамицину (Km r ) в SNC1. Смысловые и антисмысловые праймеры, соответствующие 5′- и 3′-концевым областям slr1369, соответственно, использовали для ПЦР с геномной ДНК дикого типа или SNC1 в качестве матрицы. ( c ) PG-требующий фенотип SNC1. Клетки дикого типа и SNC1 культивировали на чашках с агаром BG-11 с добавлением PG или без него.
Рост и липидные фенотипы мутанта.
SNC1 был летальным при фотоавтотрофном культивировании на агаровой пластине с BG-11, но мог расти с добавлением PG (рис. 1 c ), что указывает на то, что PG важен для цианобактерий, но не продуцируется SNC1, по крайней мере, не до уровня, поддерживающего его рост. Количественная потребность в PG для SNC1 была затем исследована путем культивирования SNC1 в жидкой среде, содержащей 0, 1,5, 6, 30 или 150 мкМ PG. Что касается скорости роста и плотности клеток в стационарной фазе, аналогичные кривые роста наблюдались с 6, 30 и 150 мкМ PG, тогда как пониженные кривые наблюдались с 0 или 1.5 мкМ PG (данные не показаны). После этого мы добавили 60 мкМ PG в среду для культивирования клеток SNC1.
Затем определяли содержание отдельных классов липидов для дикого типа и SNC1, выращенных в присутствии или в отсутствие липосом PG (рис. 2). Небольшой эффект от добавления PG наблюдался на индивидуальное содержание липидов дикого типа — то есть MGDG был наиболее распространенным липидом, за ним следовали SQDG, DGDG и PG в этом порядке (рис. 2). Соответствующее содержание MGDG, DGDG и SQDG в SNC1, выращенном с липосомами PG, было сходным с таковым в липосомах дикого типа, что указывает на то, что SNC1 мало влияет на метаболизм гликолипидов, по крайней мере, когда PG поставляется экзогенно, тогда как содержание PG в SNC1 , начиная с 0.От 1 до 5 раз выше уровня дикого типа (данные не показаны), в среднем примерно в два раза выше, чем у дикого типа (фиг. 2). Широкий диапазон содержания PG в SNC1 может зависеть от различных уровней включенного PG на фоне нарушения синтеза PG. Чтобы исследовать возможное отсутствие синтеза PG с помощью SNC1, мы переместили клетки SNC1, заранее выращенные с добавлением липосом PG, в среду, лишенную PG, а затем наблюдали за изменениями клеточного содержания PG (рис. 3). ). SNC1 показал более медленную скорость роста и более низкую плотность клеток в стационарной фазе, чем у дикого типа (рис.3 Inset ), что совпадает с требованием PG для SNC1 для роста. Содержание PG, которое оставалось на почти постоянном уровне у дикого типа на протяжении всего роста, снижалось в SNC1 с ростом в отсутствие PG. Эти результаты показывают, что SNC1 нарушает синтез PG из-за делеции гена CDS. Содержание трех других гликолипидов в SNC1 было мало изменено (данные не показаны), что указывает на то, что на метаболизм гликолипидов мало влияет дефект метаболизма PG у Synechocystis sp.PCC6803.
Рисунок 2Индивидуальный липидный состав клеток дикого типа и клеток SNC1. Индивидуальное содержание липидов определяли на основе клеток дикого типа, выращенных с добавлением PG или без него, и SNC1, выращенного с добавлением PG. Данные представлены в виде средних значений ± стандартная ошибка для трех и пяти экспериментов с диким типом и SNC1 соответственно. □, дикий тип без PG; , дикий тип с PG; ■, SNC1 с PG.
Рисунок 3Дефект PG синтеза SNC1.Клетки дикого типа выращивали в отсутствие PG, в то время как SNC1, предварительно выращенный с добавлением PG, переносили в среду без PG для анализа влияния PG на содержание в клетках. Данные представлены как средние для трех измерений в одном эксперименте, представляющем три независимых эксперимента. ( Врезка ) Кривая роста. ○, Дикий тип; ●, SNC1.
Было показано, что несколько фотосинтезирующих организмов при различных обстоятельствах демонстрируют обратные изменения уровней PG и SQDG, что приводит к небольшому изменению уровня суммы этих анионных липидов: рост вызывал снижение уровня PG с одновременным увеличением уровня SQDG у Rhodobacter sphaeroides , Synechococcus sp.PCC7942 и A. thaliana (30–32). ( ii ) Мутации, нарушающие синтез SQDG, увеличивают уровни PG в R. sphaeroides , Synechococcus sp. PCC7942 и C. reinhardtii (2, 30, 31). Таким образом, считалось, что фотосинтетическим организмам требуется определенный суммарный уровень SQDG и PG, и что каждый из этих анионных липидов в некоторой степени играет общие роли в фотосинтетическом аппарате, который может быть взят на себя, если необходимо, другим. .В отличие от приведенных выше результатов, уровень SQDG в SNC1 оставался почти постоянным, несмотря на различные уровни включения экзогенно дополненного PG (рис. 3) или снижение содержания PG после перехода на среду без PG (данные не показаны). . Одним из возможных объяснений этого несоответствия может быть то, что включенный PG в SNC1 был локализован в сайтах, которые, хотя и были достаточными для некоторого восстановления подавленного роста, были неподходящими для регуляции уровней анионных липидов. Незаменимость PG для роста Synechocystis sp.PCC6803 и тот факт, что PG не исчезал, а оставался на более низком уровне при ограничении фосфора (30–32), указывают на некоторую существенную роль PG, которая не может быть заменена SQDG.
Влияние мутации CDS на фотосинтетические пигменты.
Для определения функции PG в фотосинтетическом аппарате, SNC1, выращенный с добавкой PG, был перенесен в среду без PG, и были исследованы эффекты последующего снижения содержания PG на фотосинтетические пигменты и активность.Клеточное содержание хлорофилла (Chl) a , единственного вида Chl в комплексах PSI и PSII, оставалось почти постоянным во время роста у дикого типа, тогда как оно снижалось в SNC1 до 40% от уровня в диком типе (рис. 4 a ). Эти результаты показывают, что уровни комплексов Chl-белок снижались в клетках SNC1 с уменьшением содержания PG.
Рисунок 4Изменения содержания Chl и спектров поглощения SNC1 после перехода на среду без PG. ( a ) Клетки дикого типа выращивали в отсутствие PG, в то время как SNC1, предварительно выращенный с добавлением PG, переносили на среду без PG.Предварительно культивированные клетки были разделены на три культуры для независимого определения содержания Хл (кружок, треугольник и квадрат). Открытые символы, дикий тип; закрашенные символы, SNC1. ( b ) Клетки SNC1 выращивали с добавлением PG, а затем переводили в среду без PG для роста до тех пор, пока содержание Chl на клеточную основу не снизилось на 45%. Спектры нормированы на 730 нм.
С другой стороны, содержание фикобилисомных пигментов, таких как фикоцианин и аллофикоцианин, было одинаковым для SNC1, выращенного с липосомами PG и липосом дикого типа, и оставалось почти на одном уровне как для SNC1, так и для дикого типа на протяжении роста в среде без PG. [e.g., фикоцианин в конечном итоге составил 103–106% и 122–125% в SNC1 и диком типе, соответственно, по сравнению с содержанием в начале культивирования в среде без PG (данные не показаны)]. Эти результаты показывают, что уровни фикобилисом оставались на определенных уровнях в клетках SNC1 и дикого типа. Фикобилисомы SNC1 будут связываться на тилакоидных мембранах, а не высвобождаться в цитоплазму, ввиду сообщения о том, что диссоциация фикобилисомов от тилакоидных мембран происходит с дестабилизацией сборки субъединиц, что вызывает снижение уровней субъединиц (33).
ПоглощениеChl клетками SNC1, что согласуется с приведенными выше результатами, уменьшилось в отношении высоты пика поглощения Chl при 678 нм к высоте пика поглощения фикоцианина при 628 нм после перехода на среду без PG (рис. 4 b ). Белковые комплексы периферических мембран, такие как фикобилисомы, субъединицы которых не интегрированы в мембраны, могут не требовать PG тилакоидных мембран для построения комплексов, в отличие от интегральных мембранных белковых комплексов, таких как комплексы Chl-белок.
Влияние мутации CDS на активность PSII.
CO 2 -зависимая скорость фотосинтеза на основе Chl снижалась во время роста у дикого типа и SNC1 (рис. 5 a ). Снижение клеточного содержания Chl a в SNC1 привело к гораздо более резкому снижению в расчете на одну клетку, т. Е. Уменьшению SNC1 на 70% по сравнению с уменьшением на 30% для дикого типа (рис. . 5 b ). Эти результаты показывают, что снижение содержания PG вызывает снижение клеточного содержания Chl a и, следовательно, фотосинтетической активности клетки.Зависимая от CO 2 фотосинтетическая активность клеток SNC1 в стационарной фазе, остававшаяся на уровне 30% от максимального уровня, по-видимому, не является прямой причиной остановки роста SNC1 после переноса в среду без PG. .
Рисунок 5Изменения фотосинтетической активности SNC1 после перехода на среду без PG. Клетки дикого типа выращивали в отсутствие PG, в то время как клетки SNC1, предварительно выращенные с добавлением PG, переводили в среду без PG. Предварительно культивированные клетки были разделены на три культуры для независимых измерений фотосинтетической активности с NaHCO 3 в качестве акцептора электронов (кружок, треугольник и квадрат) на основе Chl ( a ) или на клеточной основе ( б ).Фотосинтетическая активность на клетку ( b ) была нормализована до соответствующих максимальных значений. Открытые символы, дикий тип; закрашенные символы, SNC1.
Чтобы определить влияние снижения содержания PG на фотосинтетический аппарат более подробно, мы исследовали скорость выделения кислорода ФСII в расчете на клетку с искусственным акцептором электронов, p -бензохиноном (рис. 6). Скорость выделения кислорода ФСII по сравнению с фотосинтезом, зависимым от CO 2 , составляла 1.В 3–1,7 раза выше у дикого типа на протяжении всего роста (данные не показаны). На ранней стадии роста SNC1, когда плотность клеток увеличилась в ≈2 раза от исходного уровня, скорость выделения кислорода PSII снизилась до 27–33% от исходного уровня, хотя скорость CO 2 -зависимый фотосинтез был мало затронут, а содержание Хл в клетках снизилось только до 82–87% (данные не показаны). Следовательно, активность ФСII с использованием p -бензохинона была превзойдена скоростью CO 2 -зависимого фотосинтеза.Поскольку 2-кратное увеличение плотности клеток SNC1 соответствовало снижению клеточного содержания PG до 25% от исходного уровня (рис. 3), эти результаты показывают, что уменьшение содержания PG в такой степени достаточно, чтобы в значительной степени лишить ФСII его способности использовать p -бензохинон в качестве акцептора электронов, лишь с небольшим снижением, если оно вообще есть, в уровне и фактической активности комплекса ФСII. Неспособность PSII эффективно использовать p -бензохинон указывает на конформационное изменение сайта связывания Q B на белке D1, на которое первоначально должен действовать p -бензохинон.Дальнейший рост клеток (т. Е. Гораздо большее снижение клеточного содержания PG) сопровождался снижением скорости выделения кислорода ФСII, а также содержания Chl и скорости фотосинтеза, зависящего от CO 2 , что указывает на то, что более резкое снижение содержания PG снижает уровень и фактическую активность ФСII, вероятно, из-за нестабильности аномально конформного комплекса ФСII (рис. 6). Первоначальная флуоресценция Chl, которая была почти постоянной у дикого типа, в конце концов повысилась до 4.В 5 раз больше исходного уровня в SNC1 (данные не показаны), что свидетельствует о функциональном повреждении фотосинтетического аппарата. Деструктор гена pgsA для PGP-синтазы Synechocystis sp. PCC6803 был недавно выделен и, как и SNC1, требовал добавления PG для роста и для снижения содержания PG после удаления PG из среды (34). Мутант, аналогичный SNC1, не мог проявлять активность PSII с искусственными акцепторами электронов, такими как p -бензохинон.Кроме того, мутант был поврежден в своей системе транспорта электронов от H 2 O к метилвиологену, но не в системе от восстановленного диаминодурена до метилвиологена (т.е. активности PSI). Это выявило снижение активности ФСII.
Рисунок 6Скорости p -бензохинон-зависимый O 2 эволюция и CO 2 -зависимый фотосинтез SNC1 после перехода на среду без PG. SNC1, предварительно выращенный с добавкой PG, был переведен на среду без PG.Предварительно культивированные клетки были разделены на три культуры для независимых измерений выделения кислорода (круг, треугольник и квадрат). Открытые символы, PSII; закрашенные символы, CO 2 -зависимый фотосинтез.
Ранее было показано, чтоPG тесно связан с субъединицей D1 PSII в цианобактерии, Oscillatoria chalybea , и сайт связывания был отнесен к участку гидрофобной аминокислоты между Arg 27 и Arg 225 на основе иммунореакции пептида. фрагменты расщепленной трипсином субъединицы D1 с антителами против PG (35).Кроме того, активность частиц PSII из цианобактерий подавлялась при обработке фосфолипазой A2, которая модифицировала PG, но восстанавливалась при добавлении PG (35). Изменение конформации комплекса PSII, включая сайт связывания Q B , может быть индуцировано в SNC1 и мутанте pgsA за счет истощения PG из субъединиц PSII, таких как белок D1, вызывая нарушение функции реакционный центр PSII.
У нефотосинтезирующих организмов анионные фосфолипиды, такие как PG, по-видимому, вносят вклад в процесс локализации некоторых белков в соответствующих мембранных системах.Снижение уровней как PG, так и кардиолипина (CL) у мутанта E. coli с измененным геном pgsA (PGP-синтаза) продемонстрировало ингибирование in vivo транслокации белка внешней мембраны, PhoE, через внутреннюю оболочку. мембраны и in vitro ингибирование за счет использования его внутренних мембранных везикул (6, 36). Поскольку дефицит только CL (т.е. не PG) у нуль-мутанта CL-синтазы E. coli вызвал несколько замечательных фенотипов, кроме аномальных липидных составов (12), был сделан вывод, что PG ответственен за транслокация белков, таких как PhoE, через внутренние мембраны у E.coli . С другой стороны, было показано, что пониженное содержание PG и CL в мутанте pgsA влияет на ориентацию производных Lep, белка внутренней мембраны, что привело к заключению, что топология мембранного белка зависит от содержания этих анионных липидов (7). Кроме того, участие PG в поддержании определенного уровня мембранных белковых комплексов было предложено для белкового комплекса внутренних мембран митохондрий, такого как цитохром c оксидаза, поскольку мутант S.cerevisiae , лишенные как PG, так и CL, показали пониженный уровень белкового комплекса (9, 10), тогда как нуль-мутант гена CL-синтазы этого организма не продемонстрировал каких-либо заметных вредных фенотипов, кроме отсутствия CL (13). Поскольку известно, что отсутствие некоторых индивидуальных субъединиц комплексов PSI и PSII вызывает плейотропные эффекты на другие субъединицы, приводя к разборке этих комплексов (37), вероятно, что у Synechocystis sp. PCC6803, PG требуется для нормальной интеграции некоторых субъединиц PSI и / или комплексов PSII в тилакоидные мембраны, и что нормальное построение комплексов Chl-белок затруднено из-за отсутствия PG.Необходимы дальнейшие исследования, чтобы выяснить, как PG способствует поддержанию содержания Chl в тилакоидных мембранах и конформации комплекса PSII.
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Прямые доказательства потребности в фосфатидилглицерине в фотосистеме II фотосинтеза
- Авторские права © 2000 Американское общество физиологов растений
Реферат
Считается, что фосфатидилглицерин (PG) играет важную роль в упорядоченной сборке и структурном поддержании фотосинтетического аппарата в мембранах тилакоидов. Однако его функция в фотосинтезе остается плохо изученной. В этом исследовании мы идентифицировали ген pgsA у Synechocystis sp.PCC6803, который кодирует фосфатсинтазу PG, участвующую в биосинтезе PG. Нарушение гена pgsA позволило нам манипулировать содержанием PG в мембранах тилакоидов и исследовать функцию PG в фотосинтезе. Полученный мутант pgsA мог расти только в среде, содержащей PG, и фотосинтетическая активность мутанта pgsA резко снижалась с одновременным уменьшением содержания PG в мембранах тилакоидов, когда клетки, выращенные в присутствии PG, переносились в среду. средний без PG.Это снижение фотосинтетической активности было связано со снижением активности фотосистемы (ФС) II, но не со снижением активности ФСI. Эти данные демонстрируют, что PG важен для роста Synechocystis sp. PCC6803 и представляют собой первое прямое доказательство того, что PG играет важную роль в PSII.
Мембраны тилакоидов в хлоропластах эукариотических растений и цианобактериальных клетках являются участками первичных процессов кислородного фотосинтеза. Они в основном состоят из глицеролипидов, образующих липидные бислои, и белковых комплексов, участвующих в фотосинтетическом переносе электронов и преобразовании энергии (Siegenthaler, 1998).Липидный состав тилакоидных мембран высоко консервативен среди эукариотических растений и штаммов цианобактерий и отличается от такового других мембран, которые содержат фосфолипиды в качестве основных глицеролипидов (Block et al., 1983; Murata and Nishida, 1987; Wada and Murata, 1998). Наиболее распространенными глицеролипидами тилакоидных мембран являются гликолипиды, моногалактозилдиацилглицерин (MGDG), дигалактозилдиацилглицерин (DGDG) и сульфохиновозилдиацилглицерин (SQDG). Фосфатидилглицерин (PG) — единственный фосфолипид в мембранах тилакоидов.
Роль глицеролипидов в функции тилакоидных мембран изучалась in vitro, и важные исследования были обобщены в обзоре (Siegenthaler, 1998). Однако результаты, полученные в этих исследованиях, не предоставляют прямых доказательств роли глицеролипидов в фотосинтезе и не могут быть напрямую применены к функции глицеролипидов in vivo. Функция глицеролипидов также изучалась in vivo с использованием мутантов, имеющих различный липидный состав тилакоидных мембран.Гюлер и др. (1996) сделали нулевой мутант Synechococcus sp. PCC7942 дефицит продукции SQDG за счет нарушения гена sqdB , участвующего в биосинтезе SQDG. Обширный анализ мутанта показал, что полное отсутствие SQDG не влияет на рост и фотосинтез клеток, выращенных в оптимальных условиях. Эти результаты демонстрируют, что SQDG не важен для роста цианобактерий и фотосинтеза.
Мутант с дефицитом DGDG был выделен из Arabidopsis Dörmann et al.(1995). Содержание ДГДГ в мутанте составляло около 8% от содержания ДГДГ дикого типа. Мутант показал задержку роста, бледно-зеленые листья, снижение фотосинтетической активности и измененную ультраструктуру мембран тилакоидов. Эти результаты привели к выводу, что ДГДГ важен для фотосинтеза и для поддержания структуры тилакоидных мембран. Хотя функция SQDG и DGDG была изучена с использованием мутантов, как описано выше, функция PG и MGDG не была четко изучена in vivo, поскольку мутанты, лишенные MGDG и PG, не были доступны.Härtel et al. (1998) недавно исследовали функцию PG в фотосинтезе с использованием мутанта pho1 Arabidopsis. Считается, что этот мутант нарушает загрузку фосфата ксилемы, что приводит к сильно ограниченному накоплению фосфата в стеблях и листьях (Poirier et al., 1991). Мутант реагирует на дефицит фосфата в листьях уменьшением количества фосфолипидов, особенно PG. Показано, что содержание PG в листьях мутанта на 35-45% ниже, чем в листьях дикого типа.Несмотря на это снижение PG, не было значительных различий в росте и фотосинтетической активности между мутантом и диким типом. Результаты, полученные с мутантом pho1 , позволили предположить, что PG не важен для роста и фотосинтеза. Однако мы не можем исключить возможность того, что уменьшенного количества PG все еще достаточно для достижения основной функции.
В Escherichia coli PG синтезируется из фосфатидной кислоты (PA), которая синтезируется ацилированием глицерина-3-P.PA превращается в CDP-диацилглицерин (CDP-DG) с помощью CDP-DG-синтазы, а затем в PG-фосфат (PGP) с помощью PGP-синтазы. Последней стадией биосинтеза PG является дефосфорилирование PGP, катализируемое фосфатазой PGP. Гены всех ферментов, кроме фосфатазы PGP, были клонированы из E. coli . Известно, что PG синтезируется одним и тем же путем в хлоропластах и клетках цианобактерий (Joyard et al., 1998; Wada, Murata, 1998). Однако гены ферментов, участвующих в биосинтезе PG, не были клонированы из высших растений и цианобактерий, за исключением клонирования гена CDP-DG-синтазы из картофеля и арабидопсиса (Kopka et al., 1997). Чтобы понять функцию PG в фотосинтезе, важно клонировать гены, участвующие в биосинтезе PG, и манипулировать содержанием PG в мембранах тилакоидов. Полная нуклеотидная последовательность генома Synechocystis sp. был недавно определен (Kaneko et al., 1996). Путем идентификации тех генов в геномной последовательности, которые кодируют полипептиды, гомологичные ферментам, участвующим в биосинтезе PG в E. coli , должна быть возможность манипулировать содержанием PG в этом штамме цианобактерий путем инактивации идентифицированного гена.
В этом исследовании мы идентифицировали Synechocystis sp. ген, pgsA , который кодирует синтазу PGP, и инактивировал этот ген, чтобы создать мутант, который не может синтезировать PG. Характеристика мутанта позволила нам понять функцию PG в фотосинтезе.
РЕЗУЛЬТАТЫ
Идентификация гена
pgsA из Synechocystis sp.PGP-синтаза, участвующая в биосинтезе PG, катализирует реакцию, которая формирует PGP из CDP-DG и глицерина-3-P.Гены, кодирующие этот фермент, уже были клонированы из некоторых бактерий, таких как E. coli (Ohta et al., 1981) и Bacillus subtilis (Kontinen and Tokuda, 1995). Чтобы идентифицировать ген PGP-синтазы из Synechocystis sp. мы провели поиск в базе данных (Cyanobase, Kazusa DNA Research Institute, Kisarazu, Japan) геномной последовательности Synechocystis sp. с аминокислотной последовательностью PGP-синтазы E. coli и обнаружил ген, кодирующий полипептид, гомологичный PGP-синтазе E.coli . Сравнение выведенной аминокислотной последовательности полипептида, кодируемого геном, с последовательностью PGP-синтазы E. coli показано на фиг.1. Synechocystis sp. Ген, обозначенный pgsA , кодирует открытую рамку считывания из 537 нуклеотидов, что соответствует 179 аминокислотным остаткам с приблизительной молекулярной массой 20 кДа. Две последовательности были идентичны на 38%. Это открытие предполагает, что ген Synechocystis sp. кодирует синтазу PGP.Чтобы подтвердить, что ген pgsA из Synechocystis sp. кодирует функциональную PGP-синтазу. Сначала мы экспрессировали ген в мутанте E. coli YA5512, который дефицит по PGP-синтазе (Asai et al., 1989), а затем проверили, комплементарен ли ген мутанту. Этот мутант E. coli имеет замену на одно основание в кодирующей области гена pgsA . Эта мутация изменяет аминокислотный остаток Thr-60 на Pro-60 в PGP-синтазе (Usui et al., 1994) и вызывает очень низкую активность PGP-синтазы и очень низкое содержание PG в мутанте. Содержание кардиолипина (CL) в мутанте также очень низкое, поскольку он синтезируется из PG. Мутант YA5512 растет в среде Лурия-Бертани (LB) с высокой концентрацией NaCl, но не в среде питательного бульона (NB) (Inoue et al., 1998). Этот мутант трансформировали pKK233–2 и pKK- pgsA и проверяли рост на чашках с агаром LB и NB. Подобно мутанту YA5512, трансформант YA5512 с pKK233–2 (контроль) мог расти на чашках с агаром LB, но не на чашках с агаром NB.Напротив, трансформант YA5512 с pKK- pgsA мог расти на обоих типах чашек с агаром. Это открытие демонстрирует, что мутант был дополнен геном pgsA из Synechocystis sp. относительно роста на чашке с агаром NB.
Рис.1.Сравнение вычисленной аминокислотной последовательности PGP-синтазы Synechocystis sp. кодируется геном pgsA и выведенной аминокислотной последовательностью PGP-синтазы E.coli . Аминокислотные остатки, консервативные в обеих последовательностях, обозначены звездочками. Дефисы указывают на пробелы, которые были добавлены для максимального выравнивания последовательностей.
Общие липиды, экстрагированные из трансформантов YA5512 с pKK233–2 и pKK- . PgsA анализировали с помощью тонкослойной хроматографии (ТСХ), чтобы продемонстрировать, что активность PGP-синтазы также дополнялась трансформацией геном pgsA из Synechocystis sp. В YA5512 и трансформанте с pKK233-2 были обнаружены низкие количества PG и CL (данные не показаны).Однако в трансформанте с pKK- pgsA были обнаружены высокие количества PG и CL (данные не показаны). Таблица I показывает липидный состав YA5512 и трансформантов с pKK233–2 и pKK- pgsA . Липиды, выделенные на пластине для ТСХ, анализировали с помощью газовой хроматографии. Относительное содержание PG составляло 2 мол.% В каждом из YA5512 и трансформанте с pKK233-2, тогда как содержание PG в трансформанте с pKK- pgsA было заметно увеличено до 41 мол.%. Эти результаты ясно демонстрируют, что ген pgsA из Synechocystis sp.структурный ген PGP-синтазы.
Таблица I.Липидный состав YA5512 и трансформантов YA5512, полученных с векторами экспрессии pKK233-2 и pKK-pgsA
Инактивация гена
pgsA в Synechocystis sp.Чтобы понять функцию PG в фотосинтезе, мы инактивировали ген pgsA из Synechocystis sp. путем вставки кассеты гена устойчивости к канамицину (Km r ) в кодирующую область гена pgsA , как показано на рисунке 2A.Поскольку цианобактериальные клетки обычно содержат множество копий хромосомной ДНК (Herdman et al., 1979), были проведены ПЦР и анализ гибридизации по Саузерну, чтобы подтвердить, все ли копии гена pgsA были инактивированы в мутантном штамме. На рис. 2В показаны результаты анализа ПЦР. В случае дикого типа амплифицировали фрагмент ДНК размером 0,3 т.п.н., соответствующий части нативного гена pgsA . В случае мутантного штамма амплифицировали фрагмент ДНК размером 1,5 т.п.н., соответствующий инактивированному гену pgsA , но 0.Фрагмент ДНК размером 3 т.п.н., соответствующий природному гену pgsA , не амплифицировали. Эти результаты предполагают, что все копии гена pgsA были инактивированы в мутантном штамме. Инактивация гена pgsA в мутантном штамме была также подтверждена геномным анализом гибридизации по Саузерну с использованием нативного гена pgsA в качестве ДНК-зонда (фиг. 2C). У дикого типа одиночный сигнал гибридизации был обнаружен в положении 5,4 т.п.н., тогда как в случае мутантного штамма сигнал гибридизации был обнаружен в положении 6.Положение 6 т.п.н., но сигнал в положении 5,4 т.п.н. не обнаружен. Эти результаты дополнительно демонстрируют, что все копии гена pgsA были полностью инактивированы в мутантном штамме.
Рис.2.Инсерционный мутагенез гена pgsA из Synechocystis sp. А. Структура гена pgsA дикого типа и инсерционного мутанта Synechocystis sp. Направления транскрипции гена Km r и pgsA показаны стрелками.B, ПЦР-анализ гена pgsA дикого типа (дорожка 1) и мутанта pgsA (дорожка 2). Справа указаны положения маркеров размера ДНК. Положения праймеров, используемых для реакций ПЦР, показаны стрелками на фиг. 2А. C — анализ гибридизации по Саузерну гена pgsA дикого типа (дорожка 1) и мутанта pgsA (дорожка 2). Геномные ДНК, выделенные из клеток дикого типа и мутантных клеток pgsA , переваривали Xba I.На каждую дорожку наносили один микрограмм ДНК. Мембрану гибридизовали при 42 ° C с использованием гена pgsA из Synechocystis sp. как ДНК-зонд.
Рост мутанта
pgsAРост дикого типа и мутанта pgsA в условиях фотоавтотрофного роста исследовали, чтобы понять функцию PG для роста. На фигуре 3А показан рост мутанта pgsA дикого типа на чашках с агаром. Хотя клетки дикого типа росли независимо от присутствия или отсутствия PG, мутант pgsA мог расти только на агаровой пластине, содержащей PG.
Рис.3.Профиль роста дикого типа и мутанта pgsA из Synechocystis sp. А. Рост дикого типа и мутанта pgsA на чашках с агаром. Клетки, выращенные в среде, содержащей 60 мкм PG, наносили штрихами на планшеты, содержащие 20 мкг / мл -1 канамицина и 60 мкм PG (+ PG) или без PG (-PG), и планшеты инкубировали при 30 ° C в течение 3 дней. недель. B, клетки дикого типа (○) и мутантные клетки (●) выращивали в среде, содержащей 60 мкМ PG. Значения представляют собой средние значения и sd из трех независимых экспериментов.C. Профиль роста дикого типа (○) и мутанта pgsA (●) в среде без PG. Клетки, выросшие до стационарной фазы в среде, содержащей PG, переносили в среду, не содержащую PG. Значения представляют собой средние значения и sd из трех независимых экспериментов.
Также проверяли профили роста дикого типа и мутанта pgsA в жидкой среде для выращивания. В ростовой среде, содержащей PG, идентичные кривые были получены для дикого типа и мутанта pgsA (рис.3Б). Однако, когда клетки, выращенные в среде для выращивания, содержащей PG, были перенесены в среду без PG (фиг. 3C), наблюдалась четкая разница между диким типом и мутантом. Дикий тип рос так же, как и в среде, содержащей PG, тогда как мутантный штамм pgsA прекратил рост на более ранней стадии. На фиг.4 показано влияние концентрации PG на рост мутанта pgsA . Мутантные клетки pgsA , выращенные в среде BG-11, содержащей PG, переносили в среду без PG и инкубировали в течение 3 дней, затем снова переносили в новую среду для выращивания, содержащую различные концентрации PG.Рост мутанта pgsA зависел от концентрации PG в среде для выращивания. Хотя мутант pgsA прекращал постепенно расти после инокуляции в среду, не содержащую PG, рост мутанта ускорялся в соответствии с повышенной концентрацией PG. Добавление 20 мкм PG к среде для выращивания полностью поддерживало рост мутанта. Эти результаты демонстрируют, что PG важен для роста Synechocystis sp.
Рис.4.Влияние концентрации PG на рост мутанта pgsA из Synechocystis sp. Мутантные клетки pgsA , выросшие до логарифмической фазы роста в среде, содержащей PG, переносили в среду, содержащую различные концентрации PG. ●, 0 мкм; ■, 2 мкм; ◊, 5 мкм; x, 20 мкм; ▵, 60 мкм. Значения являются средними из двух независимых экспериментов.
Чтобы изучить, могут ли другие фосфолипиды поддерживать рост мутанта pgsA , мы дополнительно проверили влияние фосфатидил-Ser, фосфатидилэтаноламина, фосфатидилхолина, PA и CL на рост этого мутанта.Однако ни один из этих фосфолипидов не был способен поддерживать рост мутанта pgsA ни в жидкой питательной среде, ни на чашках с агаром (данные не показаны). Эти результаты демонстрируют, что PG выполняет специфическую функцию, необходимую для роста Synechocystis sp.
Изменения в содержании PG после лишения PG
Для проверки изменений содержания PG в мутантных клетках pgsA после лишения PG из ростовой среды липиды были извлечены из интактных клеток и проанализированы.На фиг.5А показаны изменения содержания PG в интактных клетках дикого типа и мутанте pgsA . Хотя содержание PG у дикого типа и мутанта pgsA было по существу одинаковым, когда клетки выращивали в присутствии PG, четкое различие между двумя штаммами наблюдалось в содержании PG после лишения PG из питательная среда. Содержание PG в клетках дикого типа снизилось с 3,3 до 1,4 нмоль (10 8 клеток) -1 после депривации PG в течение 3 дней.Напротив, содержание PG в мутантных клетках pgsA заметно снизилось с 3,0 до 0,3 нмоль (10 8 клеток) -1 . Эти результаты предполагают, что мутант pgsA не способен синтезировать PG и что ген pgsA участвует в биосинтезе PG.
Рис.5.Изменения содержания PG и фотосинтетической активности интактных клеток дикого типа и мутанта pgsA из Synechocystis sp. после лишения ПГ из питательной среды.A, Изменения в содержании PG. Клетки дикого типа (○) и мутантные (●) клетки, выращенные в среде, содержащей 20 мкМ PG, переносили в среду без PG и инкубировали в течение заданного времени. Значения являются средними из двух независимых экспериментов. B, Изменения фотосинтетической активности. Клетки дикого типа (●) и мутантные (○) клетки, выращенные в среде, содержащей 20 мкМ PG, переносили в среду без PG и инкубировали в течение заданного времени. В случае мутанта культура была разделена на две колбы после лишения PG в течение 6 дней.Культуру в одной колбе дополнительно инкубировали без PG, тогда как в другую колбу добавляли 20 мкМ PG. Стрелка указывает время добавления PG. Значения являются средними из двух независимых экспериментов.
Чтобы выяснить влияние дефицита PG на состав липидов в тилакоидных мембранах, тилакоидные мембраны были проанализированы после лишения PG из ростовой среды. В таблице II показаны изменения липидного состава тилакоидных мембран, выделенных из дикого типа и мутанта pgsA .У дикого типа содержание PG снизилось с 34% до 19%, тогда как у мутанта pgsA оно уменьшилось более резко — с 15% до 4%. Содержание SQDG и MGDG незначительно увеличивалось у обоих штаммов одновременно с уменьшением содержания PG. Аналогичные изменения липидного состава наблюдались также в интактных клетках как дикого типа, так и мутанта pgsA (данные не показаны).
Таблица II.Изменения липидного состава тилакоидных мембран дикого типа и мутанта pgsA Synechocystis sp.PCC6803 после лишения PG из питательной среды
Изменения в фотосинтетической активности после лишения PG
Чтобы выяснить влияние пониженного содержания PG на фотосинтез, была измерена фотосинтетическая активность по выделению кислорода (чистый фотосинтез, вода в углекислый газ) интактных клеток дикого типа и мутанта pgsA , как показано на рисунке 5B. Фотосинтетическая активность интактных клеток дикого типа несколько увеличивалась, и эти клетки нормально росли при переносе в среду без PG.Напротив, активность интактных клеток мутанта pgsA резко снизилась после того, как эти клетки были перенесены в среду без PG. После инкубации клеток в течение 3 дней активность снизилась до 60% интактных клеток, выросших в среде, содержащей PG, и клетки перестали расти. Эта пониженная активность полностью восстанавливалась до исходного уровня активности клеток после добавления 20 мкМ PG в среду для выращивания. Этот результат предполагает, что экзогенно добавленный PG поглощался клетками и включался в мембраны тилакоидов.Включение PG в тилакоидные мембраны было подтверждено анализом липидного состава тилакоидных мембран, как описано ранее (см. Таблицу II). Это индуцированное PG восстановление активности сохранялось даже после инкубации в течение 6 дней. После восстановления фотосинтетической активности рост клеток возобновился, что свидетельствует о том, что клетки продолжали выживать даже после инкубации в течение 6 дней. Эти результаты убедительно подтверждают идею о том, что PG играет важную роль в фотосинтезе и росте Synechocystis sp.
Мы также исследовали влияние пониженного содержания PG на активность PSII. Активность PSII в интактных клетках дикого типа и мутанте pgsA была по существу одинаковой, когда клетки выращивали в присутствии PG. Интересно, что мы обнаружили, что 1,4-бензохинон (pBQ), используемый в качестве акцептора электронов от PSII, ингибировал активность мутанта pgsA после лишения PG в течение 5 дней, но не активность в интактных клетках дикой природы. тип. Как показано в таблице III, активность PSII в интактных клетках дикого типа увеличивалась при добавлении pBQ, тогда как активность в интактных клетках мутанта pgsA полностью подавлялась после лишения PG.Другие хиноны, 2,6-дихлор- p -бензохинон (DCBQ) и 2,6-диметил- p -бензохинон (DMBQ), также ингибировали активность мутантных клеток pgsA , выращенных в отсутствие PG , но с другой концентрационной зависимостью (данные не показаны). Известно, что каждый из этих хинонов проявляет различное сродство к акцепту электронов из сайта Q A белка D2 или сайта Q B белка D1 в реакционном центре PSII (Nakane et al., 1991; Satoh et al. , 1995). Эти данные свидетельствуют о том, что изменения конформации реакционного центра ФСII индуцируются снижением содержания PG в мембранах тилакоидов, и что тем самым изменяется сродство хинонов к сайту Q A или Q B .
Таблица III.Ингибирование активности PSII хинонами в интактных клетках дикого типа и мутанте pgsA Synechocystis sp. PCC6803
Чтобы дополнительно прояснить функцию PG в фотосинтезе, тилакоидные мембраны были выделены из клеток дикого типа и мутантных клеток pgsA до или после лишения PG из ростовой среды, и была измерена фотосинтетическая активность.Активности PSII и PSI тилакоидных мембран, выделенных из клеток дикого типа и мутантных клеток pgsA , сравниваются в таблице IV. Активность PSI тилакоидных мембран, измеренная с использованием восстановленного диаминодурена (DADH 2 ) и метилвиологена (MV) в качестве донора и акцептора электронов, соответственно, не подвергалась значительному влиянию депривации ни у дикого типа, ни у мутанта pgsA . PG из питательной среды. Фотосинтетическая активность тилакоидных мембран, которую измеряли как поглощение кислорода, представляющее активность переноса электронов от воды к MV, также не изменялась у дикого типа за счет лишения PG из ростовой среды.Напротив, фотосинтетическая активность от воды к МВ тилакоидных мембран мутантных клеток pgsA , выращенных в течение 6 дней без PG, снизилась примерно до 50% от активности тилакоидных мембран, выделенных из клеток, выращенных в присутствии PG. Активность ФСII тилакоидных мембран измеряли с использованием pBQ в качестве акцептора электронов. Активность тилакоидных мембран дикого типа была высокой и не изменялась при депривации PG. У мутанта pgsA активность тилакоидных мембран, выделенных из клеток, выращенных в присутствии PG, была аналогична активности тилакоидных мембран дикого типа, но активность тилакоидных мембран, выделенных после депривации PG, подавлялась. , поскольку он был в интактных клетках и не мог быть измерен путем добавления искусственных акцепторов хинона.Хотя мы не смогли измерить активность PSII тилакоидных мембран, выделенных из мутантных клеток pgsA после депривации PG, одновременно измеряемая активность транспорта электронов из воды в MV, но не активность PSI, снизилась. Эти результаты демонстрируют, что снижение содержания PG в мембранах тилакоидов после депривации PG приводит к снижению активности PSII.
Таблица IV.Три параметра фотосинтетической активности были измерены в тилакоидных мембранах, выделенных из дикого типа и мутанта pgsA Synechocystis sp.PCC6803, выращенные в присутствии или в отсутствие PG
ОБСУЖДЕНИЕ
В этом исследовании мы идентифицировали ген pgsA у Synechocystis sp. и продемонстрировали, что он кодирует PGP-синтазу на основании сходства последовательностей с PGP-синтазой E. coli и его функциональной комплементации с мутантом E. coli pgsA , YA5512, дефектным по PGP-синтазе. Идентификация гена pgsA позволила нам создать мутант Synechocystis sp.дефектный в биосинтезе PG путем инактивации гена pgsA и для изучения функции PG в фотосинтезе.
Функция PG в фотосинтезе ранее изучалась in vitro. Jordan et al. (1983) и Siegenthaler et al. (1987) сообщили, что от 70% до 80% истощения PG из тилакоидных мембран в результате переваривания мембран фосфолипазой A 2 ингибируют активность PSII и активность транспорта электронов всей цепи более чем на 50%. PG связан с белком D1 в реакционном центре PSII, и он может быть функциональным эффектором и мембранным якорем белка D1 в центральном комплексе PSII (Kruse and Schmid, 1995).Недавнее исследование также подтвердило, что PG участвует в димеризации гетеродимерных белков в комплексе PSII (Kruse et al., 2000). Хорошо известно, что скорость оборота белка D1 очень высока по сравнению с таковыми других белков (Aro et al., 1990, 1993). Эти находки предполагают, что PG играет важную роль в поддержании структуры реакционного центра PSII в тилакоидных мембранах. Однако эти исследования не смогли предоставить прямых доказательств того, что PG играет такую роль in vivo. В этом исследовании мы создали мутант pgsA из Synechocystis sp.это было неспособно синтезировать PG. Используя этот мутант, мы манипулировали содержанием PG в мембранах тилакоидов и показали, что PG необходима для роста и что активность PSII, но не активность PSI, снизилась у мутанта pgsA после лишения PG из мембран тилакоидов. Эти результаты показывают, что PG важен для роста Synechocystis sp. и играет важную роль в PSII.
Как было исследовано в настоящем исследовании, мы также разрушили ген cdsA из Synechocystis sp.которые могут кодировать синтазу CDP-DG, участвующую в биосинтезе PG, и мы исследовали функцию PG. Полученные экспериментальные данные привели нас к аналогичному выводу о важности PG для роста и активности фотосинтеза ФСII у Synechocystis sp. (Н. Сато, М. Хаджио, Х. Вада и М. Цузуки, данные представлены для публикации).
У мутанта pgsA из Synechocystis sp. Активность фотосинтеза снижалась, и клетки перестали расти при переносе в среду без PG.Вероятно, что это прекращение роста было вызвано не просто снижением активности фотосинтеза, поскольку клетки все еще демонстрировали около 60% исходной активности фотосинтеза даже после того, как клетки перестали расти. Это открытие предполагает, что PG играет важную роль не только в фотосинтезе, но и в нефотосинтетических процессах. По этой причине мы затем исследовали рост мутанта pgsA в условиях роста, при которых рост клеток не зависел от фотосинтеза.Известно, что Synechocystis sp. могут расти в условиях активированного светом гетеротрофного роста (LAHG), при котором культуры инкубируются в темноте, за исключением одного раза в день в течение 5 минут, когда они освещаются (Anderson and McIntosh, 1991). Рост клеток под действием LAHG зависит от Glc в среде для выращивания, но не от фотосинтеза. В настоящем исследовании мы исследовали рост мутанта pgsA в условиях LAHG, чтобы выяснить функцию PG в нефотосинтетических процессах.Мутант мог расти в условиях в присутствии PG, но не в отсутствие PG (данные не показаны). Это открытие ясно демонстрирует, что PG необходим даже для роста, не зависящего от фотосинтеза. Дальнейшие эксперименты с мутантом pgsA могут позволить нам определить, почему PG необходим для независимого от фотосинтеза роста Synechocystis sp.
МАТЕРИАЛЫ И МЕТОДЫ
Организмы и условия культивирования
Дикий тип Synechocystis sp.PCC6803 выращивали фотоавтотрофно при 28 ° C в среде BG-11 (Allen, 1968) с добавлением 4 мМ HEPES (2- [4- (2-гидроксиэтил) -1-пиперазинил] этансульфоновая кислота) -NaOH, pH 7,5. Мутант pgsA выращивали в той же среде BG-11, но в присутствии 20 мкг мл -1 канамицина и 20 или 60 мкм диолеоил-PG (P9664, Sigma, St. Louis). Для роста клеток на чашках с агаром в среду для роста добавляли 1,2% (мас. / Об.) Агара. Свет обеспечивался люминесцентными лампами с плотностью потока фотонов приблизительно 30 мкмоль фотонов м -2 с -1 .Культуру аэрировали на ротационном шейкере (NR-3, TAITEC, Сайтама, Япония) при 120 ударах мин. -1 . Рост культур контролировали путем определения оптической плотности при 730 нм (OD 730 ).
Комплементация мутанта
Escherichia coli YA5512 геном pgsA гена Synechocystis sp.E. coli , мутант YA5512, дефектный по PGP-синтазе, был предоставлен К. Мацумото (Университет Сайтамы).Кодирующая область гена pgsA из Synechocystis sp. амплифицировали с помощью ПЦР с двумя праймерами, 5′-CGCCATGGATATACCCAACTGGGTA-3 ‘и 5′-CGCTGCAGTTAGCCAGCCTCGACTT-3’. Восьминуклеотидные последовательности 5′-CGCCATGG-3 ‘, включая сайт Nco I и 5′-CGCTGCAG-3′, включая сайт Pst I, были добавлены к 5′-концам прямого праймера и обратного праймера. , соответственно. Амплифицированный фрагмент ДНК расщепляли Nco I и Pst I и лигировали в вектор экспрессии pKK233–2 (CLONTECH, Пало-Альто, Калифорния).Полученную плазмиду, обозначенную pKK- pgsA , использовали для трансформации E. coli YA5512. YA5512 также трансформировали pKK233–2, и полученный трансформант использовали в качестве контроля. Трансформанты YA5512, созданные с использованием экспрессионных векторов pKK- , pgsA и pKK233–2, выращивали при 37 ° C в среде LB с добавлением 100 мкг мл -1 ампициллина. Когда OD 600 культур E. coli достигла 0,5, добавляли изопропил-1-тио-β-галактозид до конечной концентрации 0.4 мм, и культуры дополнительно инкубировали в течение 3 ч при 37 ° C. После инкубации клетки собирали центрифугированием и использовали для анализа липидов.
Нарушение гена
pgsA в Synechocystis sp.Ген pgsA из Synechocystis sp. амплифицировали с помощью ПЦР с праймерами 5′-CCAAGCTTTGAACGTGGCCTTAATT-3 ‘и 5′-TACCGAGCTATCTTGGCATGATTAC-3’. Амплифицированный фрагмент ДНК лигировали в вектор pCRII (Original TA Cloning Kit, Invitrogen, San Diego).Полученную плазмиду расщепляли Eco RI с получением фрагмента ДНК размером 2,5 т.п.н., включающего ген pgsA . Фрагмент ДНК размером 2,5 т.п.н. лигировали в сайт Eco RI pBluescript II KS (+) (Stratagene, La Jolla, CA) для конструирования плазмиды pBlue- pgsA . Нарушение гена pgsA осуществляли путем вставки картриджа Km r в сайт Hpa I с тупым концом в кодирующей области гена pgsA в pBlue- pgsA .Картридж Km r получали из pUC4KIXX (Amersham, Buckinghamshire, UK) в виде фрагмента размером 1,2 т.п.н. путем расщепления с помощью Sma I. Полученная плазмида, включающая разрушенный ген pgsA (pBlue- pgsA :: Km r ) использовали для трансформации дикого типа Synechocystis sp. Трансформацию проводили, как описано Golden et al. (1987). Отбирали устойчивые к канамицину трансформанты и несколько раз наносили штрихами для разделения мутантов, у которых были нарушены все копии гена pgsA в хромосоме.Нарушение гена pgsA в мутанте pgsA было подтверждено геномным анализом гибридизации по Саузерну и ПЦР с использованием двух праймеров 5′-CCAGCACTGACTGGTTGGATGGTTA-3 ‘и 5′-AACGGTGCTAACAACAAGGCGATCG-3’.
Анализ гибридизации генома по Саузерну
Геномных ДНК, выделенных из клеток дикого типа, и мутанта pgsA из Synechocystis sp. расщепляли соответствующими рестрикционными ферментами, разделяли электрофорезом на 1% (мас. / об.) агарозном геле и переносили на нейлоновую мембрану (Hybond-N +, Amersham).Мембрану использовали для гибридизации с использованием системы мечения и обнаружения ДНК (набор ECL, Amersham). Кодирующую область гена pgsA использовали в качестве зонда.
Получение тилакоидных мембран
Тилакоидные мембраны были выделены согласно Nishiyama et al. (1993) с некоторыми изменениями. Клетки собирали из 600 мл культуры с плотностью клеток 5 мкг хлорофилла (Chl) / мл -1 центрифугированием при 5000 g в течение 10 мин. Осажденные клетки промывали один раз 100 мл среды, содержащей 50 мМ HEPES-NaOH, pH 7.5 и 30 ммCaCl 2 (среда A). Эти процедуры выполнялись при 20 ° C, а следующие процедуры выполнялись при температуре от 0 ° C до 4 ° C. Клетки ресуспендировали в 13 мл среды А, содержащей 0,8 мкМ сорбита, 1 мкМ глицинбетаина и 1 мкМ 6-аминогексановой кислоты (среда В). Суспензию смешивали с равным объемом стеклянных шариков диаметром 0,1 мм, и смесь помещали в гомогенизатор (Beadbeater, Biospec Products, Bartlesville, OK). Разрушение клеток выполняли дважды в течение 30 с, в результате чего разрушалось более 80% клеток.После осаждения стеклянных шариков супернатант центрифугировали при 2000 g в течение 5 минут для удаления неразрушенных клеток. Полученный супернатант переносили в две пробирки и затем центрифугировали при 40 000 g в течение 20 мин. Гранулированные тилакоидные мембраны в одной из двух пробирок суспендировали в воде и использовали для анализа липидов. Мембраны в другой пробирке суспендировали в среде B и использовали для измерения фотосинтетической активности. Концентрацию Хл определяли методом Arnon et al.(1974).
Измерение фотосинтетической активности
Фотосинтетическая активность измерялась посредством кислородного обмена с кислородным электродом Кларка, как описано Gombos et al. (1991) и Nishiyama et al. (1993) с некоторыми изменениями. Фотосинтетическое выделение кислорода (чистый фотосинтез) отслеживали в интактных клетках без экзогенно добавленных доноров или акцепторов электронов. Интактные клетки дважды промывали средой BG-11 и суспендировали в той же среде для измерений.Фотосинтетическую активность переноса электронов тилакоидных мембран из воды в MV измеряли в 50 мМ HEPES-NaOH, pH 7,5, 1 м глицинбетаина, 0,8 м сорбита, 0,3 мМ MV и 1 мМ NaN 3 . Активность PSII интактных клеток и тилакоидных мембран измеряли по переносу электронов от воды к pBQ. Интактные клетки дважды промывали средой BG-11 и суспендировали в среде BG-11 с добавлением 1,0 мм pBQ и 1,0 ммK 3 Fe (CN) 6 для измерений (Ono and Murata, 1981).Активность тилакоидных мембран измеряли в 50 мМ HEPES-NaOH, pH 7,5, но в присутствии 1 мМ глицинбетаина и 0,8 мМ сорбита. Активность PSI в тилакоидных мембранах измеряли по переносу электронов от DADH 2 к MV. Тилакоидные мембраны суспендировали в 50 мМ HEPES-NaOH, pH 7,5, 1,0 ммдиаминодурен, 1,0 мм Na-аскорбат, 1,5 мм MV, 0,01 мм 3- (3, 4-дихлорфенил) -1,1-диметилмочевина и 2 мм KCN ( Пакраси и др., 1988). Свет от лампы накаливания в сочетании с красным оптическим фильтром (2–61, Корнинг, Корнинг, Нью-Йорк) обеспечивал все измерения фотосинтетической активности.Интенсивность освещения составляла 500 мкмоль фотонов м −2 с −1 . Концентрации интактных клеток и мембран доводили до примерно 2 и 10 мкг Chl / мл — соответственно.
ПОДТВЕРЖДЕНИЕ
Мы благодарим профессора Коджи Мацумото (Университет Сайтамы) за предоставленный мутант E. coli pgsA (YA5512).
Сноски
№1 Работа поддержана грантом на научные исследования Министерства образования, науки, спорта и культуры (грант №12640635 к H.W.). M.H. и З.Г. были поддержаны стипендиями Японского общества содействия науке.
↵ * Автор, ответственный за переписку; электронная почта wadarcb {at} mbox.nc.kyushu-u.ac.jp; факс 81–92–726–4761.
- Поступила 10 апреля 2000 г.
- Принята 9 июня 2000 г.
Гомеостаз фосфатидилглицерина в глицерин-фосфатных ауксотрофах Staphylococcus aureus | BMC Microbiology
Zhang Y-M, Rock CO: Гомеостаз мембранных липидов у бактерий.Nat Rev Microbiol. 2008, 6: 222-233. 10.1038 / nrmicro1839.
PubMed Статья Google Scholar
Кронан Дж. Э., Rock CO: Глава 3.6.4. Биосинтез мембранных липидов. Eco-Sal-Escherichia coli и Salmonella typhimurium: клеточная и молекулярная биология. Под редакцией: Бёк И., Кертис Р. III, Капер Дж. Б., Карп П. Д., Нейдхардт Ф. К., Нистрем Т., Слаух Дж. М., Сквайрс К.Л., Уссери Д. 2008, Вашингтон, округ Колумбия: ASM Press, [ Online ] http: // www.ecosal.org
Google Scholar
Яо Дж., Rock CO: Синтез фосфатидной кислоты в бактериях. Biochim Biophys Acta. 1831, 2013: 495-502.
Google Scholar
Парсонс Дж. Б., Rock CO: Бактериальные липиды: метаболизм и мембранный гомеостаз. Prog Lipid Res. 2013, 52: 249-276. 10.1016 / j.plipres.2013.02.002.
PubMed CAS PubMed Central Статья Google Scholar
Heath RJ, Jackowski S, Rock CO: Подавление гуанозинтетрафосфатом синтеза жирных кислот и фосфолипидов в Escherichia coli облегчается сверхэкспрессией глицерин-3-фосфатацилтрансферазы ( plsB ). J Biol Chem. 1994, 269: 26584-26590.
PubMed CAS Google Scholar
Magnusson LU, Farewell A, Nystrom T: ppGpp: глобальный регулятор в Escherichia coli . ТИМ. 2005, 13: 236-242.
CAS Google Scholar
Фолькер Т.А., Дэвис Х.М.: Изменение специфичности и регуляции синтеза жирных кислот Escherichia coli путем экспрессии тиоэстеразы ацилацильного белка-носителя средней цепи растения. J Bacteriol. 1994, 176: 7320-7327.
PubMed CAS PubMed Central Google Scholar
Jiang P, Cronan JE: Ингибирование синтеза жирных кислот в Escherichia coli в отсутствие синтеза фосфолипидов и снятие ингибирования действием тиоэстеразы.J Bacteriol. 1994, 176: 2814-2821.
PubMed CAS PubMed Central Google Scholar
Чо Х, Кронан Дж. Э .: Неправильный экспорт периплазматического фермента нарушает регуляцию бактериального синтеза жирных кислот. J Biol Chem. 1995, 270: 4216-4219. 10.1074 / jbc.270.9.4216.
PubMed CAS Статья Google Scholar
Heath RJ, Rock CO: Регулирование удлинения жирных кислот и инициации белком-носителем ацилацила в Escherichia coli .J Biol Chem. 1996, 271: 1833-1836. 10.1074 / jbc.271.4.1833.
PubMed CAS Статья Google Scholar
Heath RJ, Rock CO: Ингибирование синтазы белка b-кетоацилацила-носителя (FabH) белком-носителем ацилацила в Escherichia coli . J Biol Chem. 1996, 271: 10996-11000. 10.1074 / jbc.271.18.10996.
PubMed CAS Статья Google Scholar
Дэвис М.С., Кронан Дж. Э .: Ингибирование ацетил-кофермента А карбоксилазы Escherichia coli с помощью ацилацильного белка-носителя. J Bacteriol. 2001, 183: 1499-1503. 10.1128 / JB.183.4.1499-1503.2001.
PubMed CAS PubMed Central Статья Google Scholar
Лу И-Дж, Чжан И-М, Граймс К.Д., Ци Дж., Ли Р.Э., Рок СО: Ацилфосфаты инициируют синтез мембранных фосфолипидов у грамположительных патогенов. Molec Cell.2006, 23: 765-772. 10.1016 / j.molcel.2006.06.030.
PubMed CAS Статья Google Scholar
Парсонс Дж. Б., Франк М. В., Субраманиан С., Саенкхэм П., Рок СО: метаболическая основа дифференциальной чувствительности грамположительных патогенов к ингибиторам синтеза жирных кислот. Proc Natl Acad Sci U S. A. 2011, 108: 15378-15383. 10.1073 / pnas.110
08.
PubMed CAS PubMed Central Статья Google Scholar
Schujman GE, Paoletti L, Grossman AD, De Mendoza D: FapR, бактериальный фактор транскрипции, участвующий в глобальной регуляции биосинтеза мембранных липидов. Dev Cell. 2003, 4: 663-672. 10.1016 / S1534-5807 (03) 00123-0.
PubMed CAS Статья Google Scholar
Schujman GE, Guerin M, Buschiazzo A, Schaeffer F, Llarrull LI, Reh G, Vila AJ, Alzari PM, De Mendoza D: Структурные основы регуляции биосинтеза липидов у грамположительных бактерий.EMBO J. 2006, 25: 4074-4083. 10.1038 / sj.emboj.7601284.
PubMed CAS PubMed Central Статья Google Scholar
Albanesi D, Reh G, Guerin ME, Schaeffer F, Debarbouille M, Buschiazzo A, Schujman GE, De Mendoza D, Alzari PM: Структурная основа прямой транскрипционной регуляции гомеостаза мембранных липидов у стафилококков Staphylococcus . PLoS Pathog. 2013, 9: e1003108-10.1371 / journal.ppat.1003108.
PubMed CAS PubMed Central Статья Google Scholar
Кронан Дж. Э., Вагелос П. Р.: Метаболизм и функция мембранных фосфолипидов Escherichia coli . Biochim Biophys Acta. 1972, 265: 25-60. 10.1016 / 0304-4157 (72)
-4.PubMed CAS Статья Google Scholar
Mindich L: Индукция лактозопермеазы Staphylococcus aureus в отсутствие синтеза глицеролипидов.Proc Natl Acad Sci U S. A. 1971, 68: 420-424. 10.1073 / pnas.68.2.420.
PubMed CAS PubMed Central Статья Google Scholar
Ray PH, White DC: Влияние депривации глицерина на метаболизм фосфолипидов ауксотрофа глицерина Staphylococcus aureus . J Bacteriol. 1972, 109: 668-677.
PubMed CAS PubMed Central Google Scholar
Mindich L: Мембранный синтез в Bacillus subtilis . II. Интеграция мембранных белков при отсутствии липидного синтеза. J Mol Biol. 1970, 49: 433-439. 10.1016 / 0022-2836 (70)
-Х.
PubMed CAS Статья Google Scholar
Миндич Л.: Мембранный синтез в Bacillus subtilis . I. Выделение и свойства штаммов, несущих мутации в метаболизме глицерина. J Mol Biol. 1970, 49: 415-432.10.1016 / 0022-2836 (70)
PubMed CAS Статья Google Scholar
Паолетти Л., Лу И-Дж., Шуйман Г.Е., Де Мендоза Д., Рок СО: Сочетание синтеза жирных кислот и фосфолипидов в Bacillus subtilis . J Bacteriol. 2007, 189: 5816-5824. 10.1128 / JB.00602-07.
PubMed CAS PubMed Central Статья Google Scholar
Крейсвирт Б.Н., Лофдал С., Бетли М.Дж., О’Рейли М., Шливерт П.М., Бергдолл М.С., Новик Р.П.: структурный ген экзотоксина синдрома токсического шока не передается через профаг. Природа (Лондон). 1983, 305: 709-712. 10.1038 / 305709a0.
CAS Статья Google Scholar
Парсонс Дж. Б., Яо Дж., Франк М. В., Джексон П., Рок КО: Разрушение мембраны антимикробными жирными кислотами высвобождает низкомолекулярные белки из Staphylococcus aureus .J Bacteriol. 2012, 194: 5294-5304. 10.1128 / JB.00743-12.
PubMed CAS PubMed Central Статья Google Scholar
Чжун Дж., Карберг М., Ламбовиц А.М.: Целевое и случайное нарушение бактериального гена с использованием интронного вектора группы II (таргетрона), содержащего селективный маркер, активируемый ретротранспозицией. Nucleic Acids Res. 2003, 31: 1656-1664. 10.1093 / нар / гкг248.
PubMed CAS PubMed Central Статья Google Scholar
Bligh EG, Dyer WJ: Быстрый метод экстракции и очистки общих липидов. Может J Biochem Physiol. 1959, 37: 911-917. 10.1139 / о59-099.
PubMed CAS Статья Google Scholar
Минклер П.Е., Кернер Дж., Ингаллс С.Т., Хоппель К.Л.: Новая процедура выделения эфиров ацилкофермента А с короткой, средней и длинной цепью из тканей. Анальная биохимия. 2008, 376: 275-276. 10.1016 / j.ab.2008.02.022.
PubMed CAS PubMed Central Статья Google Scholar
Haynes CA, Allegood JC, Sims K, Wang EW, Sullards MC, Merrill AH: Количественное определение жирного ацил-кофермента As в клетках млекопитающих с помощью тандемной масс-спектрометрии с ионизацией жидкой хроматографией и электрораспылением. J Lipid Res. 2008, 49: 1113-1125. 10.1194 / мл. D800001-JLR200.
PubMed CAS PubMed Central Статья Google Scholar
Bell RM: Мутанты Escherichia coli , дефектные по мембранному синтезу фосфолипидов: макромолекулярный синтез в мутанте sn -глицерин-3-фосфатацилтрансферазы K m .J Bacteriol. 1974, 117: 1065-1076.
PubMed CAS PubMed Central Google Scholar
Валлари Д.С., Яковски С., Рок СО: Регулирование пантотенаткиназы коферментом А и его тиоэфирами. J Biol Chem. 1987, 262: 2468-2471.
PubMed CAS Google Scholar
Grundling A, Schneewind O: Синтез глицеринфосфат липотейхоевой кислоты в Staphylococcus aureus .Proc Natl Acad Sci U S. A. 2007, 104: 8478-8483. 10.1073 / pnas.0701821104.
PubMed PubMed Central Статья Google Scholar
Jerga A, Lu Y-J, Schujman GE, De Mendoza D, Rock CO: Идентификация растворимой диацилглицерин-киназы, необходимой для производства липотейхоевой кислоты в Bacillus subtilis . J Biol Chem. 2007, 282: 21738-21745. 10.1074 / jbc.M703536200.
PubMed CAS Статья Google Scholar
Кирюхин М.Ю., Дебабов Д.В., Шинабаргер Д.Л., Neuhaus FC: Биосинтез гликолипидного якоря липотейхоевой кислоты Staphylococcus aureus RN4220: роль YpfP, диглюкозилдиацилглицеринсинтазы. J Bacteriol. 2001, 183: 3506-3514. 10.1128 / JB.183.11.3506-3514.2001.
PubMed CAS PubMed Central Статья Google Scholar
Оку Ю., Курокава К., Ичихаши Н., Секимидзу К. Характеристика гена Staphylococcus aureus mprF , участвующего в лизинилировании фосфатидилглицерина.Микробиология. 2004, 150: 45-51. 10.1099 / mic.0.26706-0.
PubMed CAS Статья Google Scholar
Koprivnjak T, Zhang D, Ernst CM, Peschel A, Nauseef WM, Weiss JP: Характеристика Staphylococcus aureus кардиолипинсинтаз 1 и 2 и их вклад в накопление кардиолипина в стационарной фазе и внутри фагоцитов. J Bacteriol. 2011, 193: 4134-4142. 10.1128 / JB.00288-11.
PubMed CAS PubMed Central Статья Google Scholar
Цай М., Охнива Р.Л., Като Ю., Такешита С.Л., Охта Т., Сайто С., Хаяси Х., Морикава К.: Золотистый стафилококк требует кардиолипина для выживания в условиях высокой солености. BMC Microbiol. 2011, 11: 13-10.1186 / 1471-2180-11-13.
PubMed CAS PubMed Central Статья Google Scholar
Охнива Р.Л., Китабаяси К., Морикава К.: Альтернативная кардиолипинсинтаза Cls1 компенсирует остановленную функцию Cls2 у Staphylococcus aureus в условиях острого кислотного стресса.FEMS Microbiol Lett. 2013, 338: 141-146. 10.1111 / 1574-6968.12037.
PubMed CAS Статья Google Scholar
(PDF) Существенная роль фосфатидилглицерина в фотосинтезе
Бенсон А.А., Маруо Б. (1958) Фосфолипиды растений. Идентификация
фосфатидилглицеринов. Biochim Biophys Acta 27: 189–195
Бенсон А.А., Маруо Б. (1989) «Нова» в метаболизме фосфатов,
GPG и открытие фосфатидилглицерина.Biochim Biophys
Acta 1000: 447–451
Block MA, Dorne A-J, Joyard J, Douce R (1983) Подготовка и
характеристика мембранных фракций, обогащенных внешней и
внутренней оболочкой мембран из хлоропластов шпината: II. Bio-
химическая характеристика. J Biol Chem 258: 13281–13286
Browse J, McCourt P, Somerville CR (1985) Мутант Arabidopsis
, лишенный специфичного для хлоропласта липида. Science 227: 763–765
Browse J, Somerville C (1991) Синтез глицеролипидов — биохимия
и регулирование.Annu Rev Plant Physiol Plant Mol Biol 42: 467–
506
Карман Г.М., Генри С.А. (1999) Биосинтез фосфолипидов в дрожжах
Saccharomyces cerevisi ae и взаимосвязь с другими
метаболическими процессами. Prog Lipid Res 38: 361–399
Demonkos I, Malec P, Sallai A, Kova
´
cs L, Itoh K, Shen G, Ughy B,
Bogos B, Sakurai I, Kis M, Strzalka K , Wada H, Itoh S, Farkas
T, Gombos Z (2004) Фосфатидилглицерин необходим для
олигомеризации реакционного центра фотосистемы I.Plant Physiol
134: 1471–1478
Дорн А.Дж., Джойард Дж., Дус Р. (1990) Действительно ли тилакоиды содержат
фосфатидилхолин? Proc Natl Acad Sci USA 87: 71–74
Dowhan W (1997) Молекулярная основа мембранного фосфолипида
Разнообразие: почему так много липидов? Annu Rev Biochem
66: 199–232
Droppa M, Horva
´
th G, Hideg E, Farkas T (1995) Роль фосфолипидов
в регулировании транспорта фотосинтетических электронов
активности: лечение тилакоидов с фосфолипазой C.Фото
synth Res 46: 287–293
Dubertret G, Mirshahi A, Mirshahi M, Gerard-Hirne C., Tremolieres
A (1994) Данные, полученные в результате манипуляций in vivo с липидным составом
у мутантов, D
3
-транс-гексадеценовая кислота, содержащая
фосфатидилглицерин, участвует в биогенезе света-
, собирающего хлорофилл a / b-белковый комплекс Chlamydomonas
reinhardtii. Eur J Biochem 226: 473–482
Dubertret G, Gerard-Hirne C, Tre
´
molie
`
res A (2002) Важность
транс-D
3
-hexadecenoic acid -содержащий фосфатидилглицерин
, образование тримерного светособирающего комплекса
Chlamydomonas.Plant Physiol Biochem 40: 829–836
Frentzen M (2004) Фосфатидилглицерин и сульфохиновозилдиацил-
глицерин: анионные мембранные липиды и регуляция фосфатов.
Curr Opin Plant Biol 7: 270–276
Frentzen M, Heinz E, McKeon TA, Stumpf PK (1983) Спецификации
и селективность глицерин-3-фосфатацилтрансферазы из
хлоропластов гороха и шпината. Eur J Biochem 129: 629–636
Frentzen M, Nishida I, Murata N (1987) Свойства пластидий
ацил- (белок-носитель ацила): глицерол-3-фосфат-ацилтрансфераза
из чувствительного к холоду растение тыквенное (Cucurbita moschata).
Physiol растительных клеток 28: 1195–1201
Gombos Z, Va
´
rkonyi Z, Hagio M, Iwaki M, Kova
´
cs L, Masamoto K,
Itoh 2002) Требование фосфатидилглицерина для функции
акцептора электронов пластохинона Q
B
в реакционном центре photosys-
tem II. Biochemistry 41: 3796–3802
Griebau R, Frentzen M (1994) Биосинтез фосфатидилглицерина
в изолированных митохондриях этиолированных проростков маша (Vigna radiata
L).Plant Physiol 105: 1269–1274
Hagio M, Gombos Z, Va
´
rkonyi Z, Masamoto K, Sato N, Tsuzuki M,
Wada H (2000) Прямые доказательства потребности в фосфате-
идилглицерине в фотосистеме II фотосинтеза. Plant Physiol
124: 795–804
Hagio M, Sakurai I, Sato S, Kato T, Tabata S, Wada H (2002)
Фосфатидилглицерин необходим для развития коидных мембран thyla-
у Arabidopsis thaliana.Physiol растительных клеток
43: 1456–1464
Hobe S, Prytulla S, Ku
¨
hlbrandt W., Paulsen H (1994) Тримеризация
и кристаллизация восстановленного светособирающего комплекса хлорофилла
a / b. EMBO J 13: 3423–3429
Hobe S, Fo
¨
rster R, Klingler J, Paulsen H (1995) N-проксимальная последовательность
мотив в собирающем свет хлорофиллом a / b-связывающем белке
необходим для тримеризации светособирающего комплекса хлорофилла a / b
.Biochemistry 34: 10224–10228
Ishizaki O, Nishida I, Agata K, Eguchi G, Murata N (1988) Клонирование
и нуклеотидной последовательности кДНК для пластид глицерин-3-
фосфатацилтрансферазы из кабачков. FEBS Lett 238: 424–430
Ishizaki-Nishizawa O, Fujii T, Azuma M, Sekiguchi K, Murata N,
Ohtani T, Toguri T (1996) Низкотемпературная стойкость высших растений
значительно повышается за счет неспецифическая цианобактериальная
десатураза.Nat Biotechnol 14: 1003–1006
Jordan BR, Chow W-S, Baker AJ (1983) Роль фосфолипидов в
молекулярной организации мембран хлоропластов гороха: влияние
истощения фосфолипидов на фотосинтетическую активность. Biochim
Biophys Acta 725: 77–86
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W., Krauß N (2001)
Трехмерная структура фотосистемы I цианобактерий на
2,5 A
˚
Разрешение.Nature 411: 909–917
Joyard J, Mare
´
chal E, Miege C, Block MA, Dorne A-J, Douce R
(1998) Структура, распределение и биосинтез глицеролипидов
из хлоропластов высших растений. В: Siegenthaler P-A, Murata N
(eds) Липиды в фотосинтезе. Kluwer Academic Publishers,
Dordrecht, Нидерланды, стр. 21–52
Kenrick JR, Bishop DG (1986) Фосфатидилглицерин и сульфохи
новосилдиацилглицерин в листьях и плодах чувствительных к холоду растений
.Phytochemistry 25: 1293–1295
Kruse O, Schmid GH (1995) Роль фосфатидилглицерина как функционального эффектора
и мембранного якоря пептида ядра D1
из частиц фотосистемы II цианобактерии Oscillato ria
чалибея. Z Naturforsch 50c: 380–390Kruse O, Hankamer B, Konczak C, Gerle C, Morris E, Radunz A,
Schmid GH, Barber J (2000) Фосфатидилглицерин участвует в
димеризации фотосистемы II.J Biol Chem 275: 6509–6514
Kurisu G, Zhang H, Smith JL, Cramer WA (2003) Структура цитохрома b
b
6
f комплекса оксигенного фотосинтеза: настройка полости
. Science 302: 1009–1014
Лю З., Янь Х., Ван К., Куанг Т., Чжан Дж., Гуй Л., Ань Х, Чанг В.
(2004) Кристаллическая структура основного светозубого жала шпината
Комплекспри 2,72 Разрешение
˚
. Nature 428: 287–292
Loll B, Kern J, Seanger W., Zouni A, Biesiadka J (2005) К
полному расположению кофакторов в 3.0 A
˚
структура разрешения
фотосистемы II. Nature 438: 1040–1044
Lyons JK (1973) Повреждение растений, вызванное переохлаждением. Annu Rev Plant Physiol
24: 445–466
Maanni AE, Dubertret G, Delrieu MJ, Roche O, Tre
´
molie
`
res A (1998)
Мутанты, пораженные хламидиозом фосфат-
метаболизм идилглицерина и биогенез тилакоидов. Plant Physiol
Biochem 36: 609–619
Малкин Р., Нийоги К. (2000) Фотосинтез.В: Buchanan BB,
Gruissem W, Jones RL (eds) Биохимия и молекулярная биология
растений. Американское общество физиологов растений,
Роквилл, Мэриленд, стр.