Содержание белка в зерновых культурах таблица: Сколько белка в кукурузе и пшенице, количество протеина в зерновых
Содержание белка и аминокислот в зерне озимых культур, произрастающих на территории лесостепи юго-востока Западной Сибири Текст научной статьи по специальности «Сельское хозяйство, лесное хозяйство, рыбное хозяйство»
DOI: 10.14258/jcprm.201503754
УДК 577.112.3:633.324 (571.1)
СОДЕРЖАНИЕ БЕЛКА И АМИНОКИСЛОТ В ЗЕРНЕ ОЗИМЫХ КУЛЬТУР, ПРОИЗРАСТАЮЩИХ НА ТЕРРИТОРИИ ЛЕСОСТЕПИ ЮГО-ВОСТОКА ЗАПАДНОЙ СИБИРИ
© Е.П. Кондратенко1, О.Б. Константинова1, О.М. Соболева1, Е.А. Ижмулкина1, Н.В. Вербицкая1, A.C. Сухих2
1 Кемеровский государственный сельскохозяйственный институт, ул. Марковцева, 5, Кемерово, 650056 (Россия), e-mail: [email protected] 2Кемеровская государственная медицинская академия, ул. Ворошилова, 22а, 650029 (Россия).
Представлены результаты анализа содержания белка и аминокислот в зерне озимой пшеницы, озимой ржи и озимого тритикале, произрастающих на территории лесостепи юго-востока Западной Сибири в зависимости от вида культуры и биологических особенностей сорта.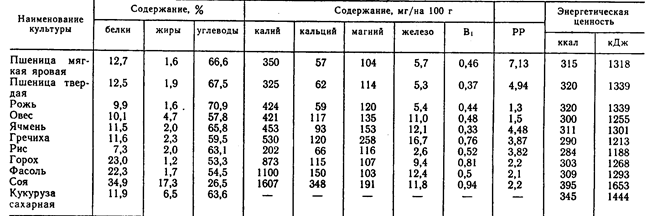
Ключевые слова: сорт, зерно, озимая пшеница, озимая рожь, озимый тритикале, белок, аминокислоты, аминокислотный скор, биологическая ценность.
Задача получения качественной, экологически чистой и сбалансированной по аминокислотному составу сельскохозяйственной продукции, в частности кормового и продовольственного зерна, в последнее
время становится актуальной.
Проблема качества зерна в современной селекции — одна из главных, наряду с продуктивностью. Решение данной задачи возможно на основе использования всех ресурсов повышения качества сельскохозяйственных культур [1]. В сложных агроклиматических условиях Сибири в сельском хозяйстве интенсивного типа все большее значение приобретают хорошо адаптированные к определенным географическим зонам сорта растений. Внедрение адаптированных сортов в сельскохозяйственное производство становится сейчас наиболее эффективным способом увеличения валовых сборов и повышения качества зерна в хозяйствах юго-востока Западной Сибири.
Решение данной задачи возможно на основе использования всех ресурсов повышения качества сельскохозяйственных культур [1]. В сложных агроклиматических условиях Сибири в сельском хозяйстве интенсивного типа все большее значение приобретают хорошо адаптированные к определенным географическим зонам сорта растений. Внедрение адаптированных сортов в сельскохозяйственное производство становится сейчас наиболее эффективным способом увеличения валовых сборов и повышения качества зерна в хозяйствах юго-востока Западной Сибири.
Кондратенко Екатерина Петровна — профессор кафедры технологии хранения и переработки сельскохозяйственной продукции, доктор сельскохозяйственных наук, e-mail: [email protected] Константинова Ольга Борисовна — аспирант кафедры технологии хранения и переработки сельскохозяйственной продукции, e-mail: [email protected]
Соболева Ольга Михайловна — доцент кафедры технологии хранения и переработки сельскохозяйственной продукции, кандидат биологических наук, e-mail: meer@yandex. ru Ижмулкина Екатерина Александровна — начальник отдела инновационных технологий, e-mail: [email protected] Вербицкая Наталья Валерьевна — аспирант кафедры технологии хранения и переработки сельскохозяйственной продукции, e-mail: [email protected] Сухих Андрей Сергеевич — старший научный сотрудник центральной научно-исследовательской лаборатории, кандидат фармацевтических наук, доцент, e-mail: Suhih [email protected]
ru Ижмулкина Екатерина Александровна — начальник отдела инновационных технологий, e-mail: [email protected] Вербицкая Наталья Валерьевна — аспирант кафедры технологии хранения и переработки сельскохозяйственной продукции, e-mail: [email protected] Сухих Андрей Сергеевич — старший научный сотрудник центральной научно-исследовательской лаборатории, кандидат фармацевтических наук, доцент, e-mail: Suhih [email protected]
Автор, с которым следует вести переписку.
Содержание белка и аминокислот — одна из важнейших характеристик биологической ценности зерна. Аминокислотный состав используется как биохимический критерий биологической ценности кормов и пищевых продуктов (по суммарному содержанию незаменимых аминокислот). Аминокислоты являются структурными единицами белковых молекул, участвующих во всех процессах, происходящих в организме человека и животных. Без белков невозможны жизнь, рост и развитие организма.
Дефицит протеинов стимулирует поиск новых сортов и гибридов с высоким содержанием протеина, с хорошо сбалансированным аминокислотным составом, которые могли бы быть использованы в селекционных программах для создания новых улучшенных сортов.
В качестве основного белоксодержащего сырья ведущая роль принадлежит зерновым злаковым культурам. В глобальном масштабе около 70% потребности человечества в белках покрывается за счет зерна — или при непосредственном употреблении в пищу, или опосредованно путем скармливания его животным для производства мяса. Следовательно, запасные белки растений служат основой питания человека.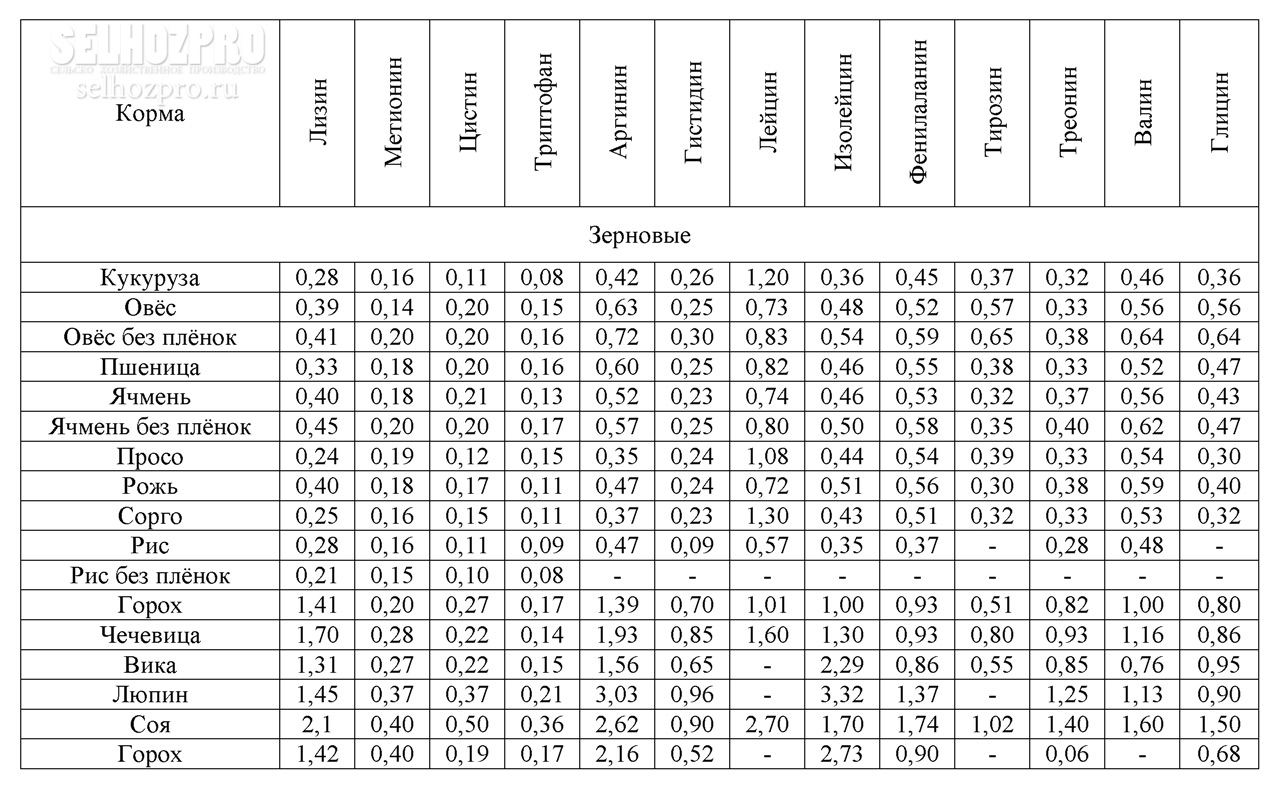
В России озимая пшеница — основная продовольственная культура, площади ее посева по годам колеблются от 8 до 11 млн га, на ее долю приходится от 20 до 24% валового сбора зерна. Озимая пшеница позволяет решить вопросы продовольственного обеспечения населения. Ее ценность состоит в том, что зерно отличается высоким содержанием белка (14-17% и более) и углеводов (80%), она широко используется в хлебопекарной, макаронной, кондитерской промышленности [2] .
Главным производителем ржи является Российская Федерация. Около 30% производства ржи используется для пищи человека, остальная часть идет на корм для животных. Основное назначение озимой ржи — продовольственное [3]. В зерне ржи, в зависимости от условий выращивания и сорта, содержится 917% белка, 52-63% крахмала, 1,6-1,9% жира [4]. Белок озимой ржи по аминокислотному составу более сбалансирован, чем пшеницы и других зерновых культур, он богаче лизином, треонином, фенилаланином и аргинином [5].
Озимый тритикале — ценная кормовая и продовольственная культура. Тритикале сочетает в себе хозяйственно-ценные признаки пшеницы и ржи, отличается повышенной устойчивостью к болезням и неблагоприятным погодным условиям, способен накапливать в зерне большое количество белка. Например, О.И. Босиева с соавт. отмечают, что тритикале на территории Республики Северная Осетия — Алания может накапливать белка до 16% [8]. В Сибири площади возделывания сортов тритикале в последние годы достигают нескольких десятков тысяч гектаров [9]. По аминокислотному составу белки этой культуры занимают промежуточное положение между белками пшеницы и ржи [10].
Положительные особенности озимых культур обусловливают стремление поиска исследований качества зерна, получаемого на юго-востоке Западной Сибири, в частности в Кемеровской области.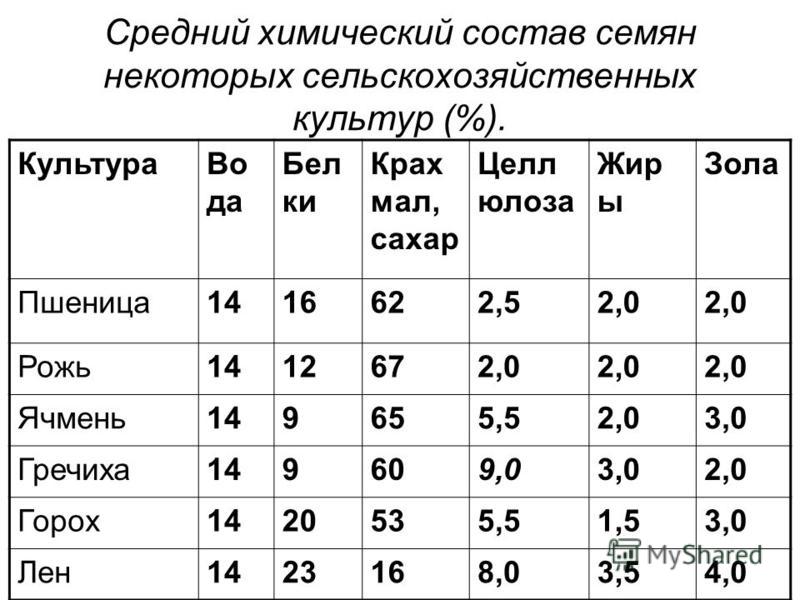 На территории области площади посева озимых зерновых злаковых культур в 2013 г. составили 23294 га. Озимые культуры представлены в основном озимой рожью (14186 га), небольшие площади (более 10000 га) занимает озимая пшеница, и в последние годы все шире распространяется сравнительно новая культура — тритикале. Доля ее посева под урожай в 2013 г. составила 3% озимого клина.
На территории области площади посева озимых зерновых злаковых культур в 2013 г. составили 23294 га. Озимые культуры представлены в основном озимой рожью (14186 га), небольшие площади (более 10000 га) занимает озимая пшеница, и в последние годы все шире распространяется сравнительно новая культура — тритикале. Доля ее посева под урожай в 2013 г. составила 3% озимого клина.
Известно, что количество аминокислот в зависимости от видовых и сортовых особенностей, а также от факторов внешней среды обеспечивает накопление отдельных фракций белковых веществ. Это в конечном итоге и определяет количество и качество формируемых белков. Нераскрытыми остаются вопросы реакции генотипа на накопление белка в зерне озимой пшеницы, озимой ржи и озимого тритикале, произрастающих в гидротермических условиях лесостепи Кемеровской области.
Цель данного исследования — изучение содержание белка и аминокислот в зерне озимой пшеницы, озимой ржи и озимого тритикале, произрастающих в условиях лесостепи юго-востока Западной Сибири.
Экспериментальная часть
Объектом исследования служили три новых сорта озимой пшеницы российской селекции — Скипетр (оригинатор Г.М. Полетаев), Новосибирская 2 и Новосибирская 3 (оригинатор — СибНИИРС), два сорта озимой ржи российской селекции — Петровна 1 (оригинатор — СибНИИСХ и торфа), сорт иностранной селекции — Зу Драйве и гибрид первого поколения (F|) — Хеллтоп, два сорта озимого тритикале — Алтайская 5 (оригинатор — Алтайский НИИ земледелия и селекции), Омская (оригинатор — СибНИИСХ и торфа). Все изучаемые сорта относятся к среднеспелой группе. Вегетационный период составляет 316-320 суток. Для оценки содержания белка (по ГОСТ 10846-91), в зависимости от биологических особенностей вида и сорта, были взяты образцы зерна разных сортов озимых злаковых зерновых культур, выращенных в одинаковых агроклиматических условиях — территория лесостепной зоны юго-востока Западной Сибири.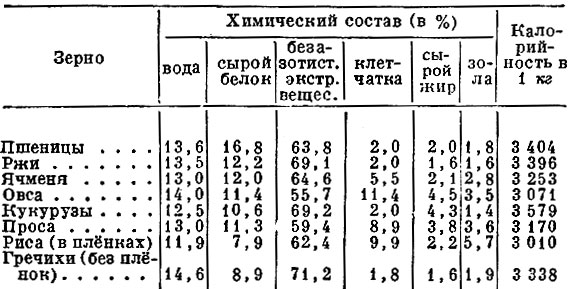
Определение аминокислотного состава зерна изучаемых сортов проведено на аминокислотном анализаторе NIR Systems 4500 (США) в лаборатории биологической химии ГНУ СибНИИЖ Россельхозакадемии (Новосибирск).
Качество белка зерна оценивали путем сравнения его аминокислотного состава с аминокислотным составом «идеального» белка с помощью расчета его аминокислотного скора, который осуществляли по формуле:
С= А /Н,
где С — скор, %;А- содержание аминокислоты в белке зерна изучаемого сорта, г/100 г белка; Н — содержание аминокислоты в идеальном белке, г/100 г белка (ФАО/ВОЗ, 1973).
Биологическую ценность белка определяли по формуле:
БЦ =100 -КРАС-,
V АРАС
КРАС = —-,
п
где ЕДРАС — разность аминокислотного скора для каждой незаменимой аминокислоты по сравнению с одной из наиболее дефицитных; п — число незаменимых аминокислот.
На основе этого показателя выделяется три уровня качества: высокий (степень отклонения от стандарта) до 15%; средний — от 16 до 30%; низкий — выше 31%, по которым делали заключение о качестве зерна [11].
Для оценки степени варьирования рассчитывали коэффициент вариации, определяющийся как отношение среднеквадратичного отклонения к среднему значению, выраженному в процентах (V, %).
Результаты и обсуждение
Анализ качества зерна показал, что более высоким содержанием белка характеризовалось зерно озимого тритикале (14,2%), меньшим — озимой ржи (10,75%) (табл. 1). Значение данного показателя варьировало в зерне озимой пшеницы от 11,30 до 13,43%, озимой ржи — от 10,14 до 11,78% и озимого тритикале-от 14,18 до 14,31%.
Проявились сортовые отличия по содержанию белка в зерне. По совокупности данного показателя более качественное зерно формировал сорт озимой пшеницы Новосибирская 2, озимой ржи — Петровна 1 и озимого тритикале — Алтайская 5 и Омская, т.е. сорта сибирской селекции (рис. 1).
По совокупности данного показателя более качественное зерно формировал сорт озимой пшеницы Новосибирская 2, озимой ржи — Петровна 1 и озимого тритикале — Алтайская 5 и Омская, т.е. сорта сибирской селекции (рис. 1).
Данные рисунка 1 наглядно демонстрируют влияние генотипа и видовых особенностей культуры на содержание белка в зерне злаковых растений, так как влияние эдафических и погодных факторов было одинаковым. Среди сортов озимой пшеницы наибольшее содержание белка отмечено у сорта Новосибирская 2 — 13,43%, что на 0,85 и 2,13 процентных пункта выше, чем у сортов Скипетр и Новосибирская 3. Среди сортов озимой ржи превосходство за сортом Петровна 1, зерно которого содержит белка больше на 1,44 и 1,64% по сравнению с сортом иностранной селекции Зу Драйве и гибридом (F|) Хеллтоп соответственно. Высоким содержанием белка характеризовались сорта озимого тритикале Алтайская 5 и Омская -14,31 и 14,08% соответственно.
Сравнительное изучение аминокислотного состава белков показало, что в среднем по содержанию всех аминокислот зерно озимой ржи уступает зерну остальных изучаемых культур. Суммарное содержание аминокислот больше в сортах озимой пшеницы и озимого тритикале на 0,84 и 1,23% соответственно (табл. 2).
Суммарное содержание аминокислот больше в сортах озимой пшеницы и озимого тритикале на 0,84 и 1,23% соответственно (табл. 2).
Таблица 1. Среднее содержание белка в зерне озимых злаковых культур за период 2012-2013 гг.
Показатель
Массовая доля, %
озимая пшеница
озимая рожь
озимыи тритикале
Среднее Lim V,%
12,41 11,30-13,43 15,86
10,75 10,14-11,78 13,92
14,2 14,08-14,31
1,6
Рис. 1,35 3,23 3,18-3,28
1,35 3,23 3,18-3,28
Пролин 0,32 0,19-0,51 0,79 0,37-1,26 0,94 0,65-1,23
Глицин 0,67 0,63-0,70 0,69 0,68-0,70 0,67 0,66-0,69
Алании 0,36 0,36-0,36 0,25 0,22-0,30 0,30 0,26-0,34
Валин 0,68 0,59-0,75 0,61 0,54-0,64 0,61 0,58-0,65
Метионин 0,31 0,27-0,34 0,11 0,06-0,15 0,24 0,19-0,30
Изолейцин 0,65 0,61-0,70 0,39 0,36-0,43 0,56 0,53-0,59
Лейцин 0,55 1,45-1,61 1,52 1,17-1,85 1,62 1,51-1,73
Тирозин 0,28 0,26-0,31 0,16 0,09-0,26 0,23 0,22-0,25
Фенил ал анин 0,68 0,64-0,74 0,34 0,18-0,59 0,57 0,31-0,83
Гистидин 0,27 0,26-0,28 0,25 0,16-0,33 0,30 0,26-0,35
Лизин 0,43 0,40-0,46 0,24 0,18-0,31 0,37 0,31-0,42
Аргинин 0,23 0,17-0,26 0,09 0,04-0,15 0,24 0,07-0,42
Триптофан 0,11 0,09-0,12 0,13 0,11-0,15 0,13 0,12-0,14
Цистин 0,24 0,19-0,29 0,22 0,20-0,24 0,18 0,17-0,20
Сумма 12,02 — 11,18 — 12,41 —
Приблизительно одинаковым в зерне озимой пшеницы, озимой ржи и озимого тритикале оказывается содержание глицина (0,67-0,69 г/100 г), аланина (0,25-0,36 г/100 г), гистидина (0,25-0,30 г/100 г), триптофана (0,11-0,13 г/100 г) и цистина (0,18-0,24 г/100 г).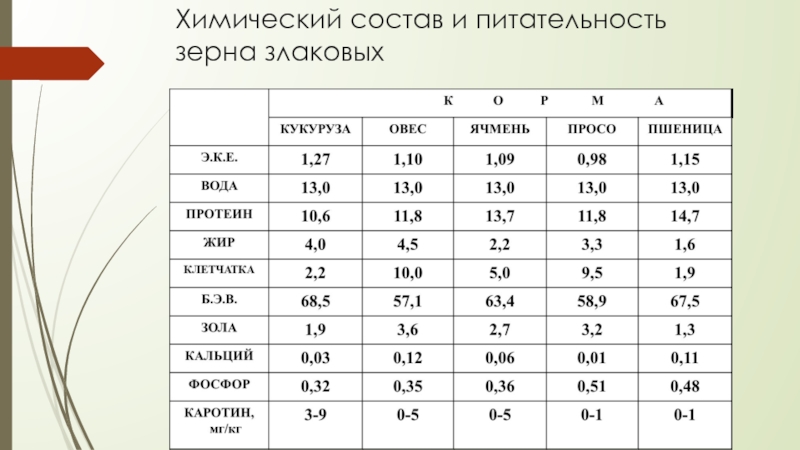 Содержание триптофана в белках всех озимых культур, произрастающих на территории юго-востока Западной Сибири, низкое. Из литературы известно [12], что эта незаменимая аминокислота является предшественником для образования фитогормона индо-лилуксусной кислоты (ИДК, indoleacetic acid, IAA) и витамина РР, роль которых в жизни растений общеизвестна. Необеспеченность семян триптофаном может привести к нарушению обмена веществ, снижению их всхожести и жизнеспособности.
Содержание триптофана в белках всех озимых культур, произрастающих на территории юго-востока Западной Сибири, низкое. Из литературы известно [12], что эта незаменимая аминокислота является предшественником для образования фитогормона индо-лилуксусной кислоты (ИДК, indoleacetic acid, IAA) и витамина РР, роль которых в жизни растений общеизвестна. Необеспеченность семян триптофаном может привести к нарушению обмена веществ, снижению их всхожести и жизнеспособности.
|4М JJ0S
10.14 мм
Озимьш тритикале
Озимая пшеница
Озимая рожь
В белках озимой ржи мало аргинина (0,09 г/100 г) и метионина (0,11 г/100 г), но больше глютаминовой кислоты. По содержанию остальных аминокислот зерно озимой пшеницы, озимой ржи и озимого тритикале существенно различается, особенно по содержанию незаменимых аминокислот — треонина (0,77; 0,63 и 0,66 г/100 г), метионина (0,31; 0,11 и 0,24 г/100 г), изолейцина (0,65; 0,39 и 0,56 г/100 г), лейцина (0,55; 1,52 и 1,62 г/100 г), фенилаланина (0,68; 0,34 и 0,57 г/10 г), лизина (0,43; 0,24 и 0,37 г/100 г) соответственно.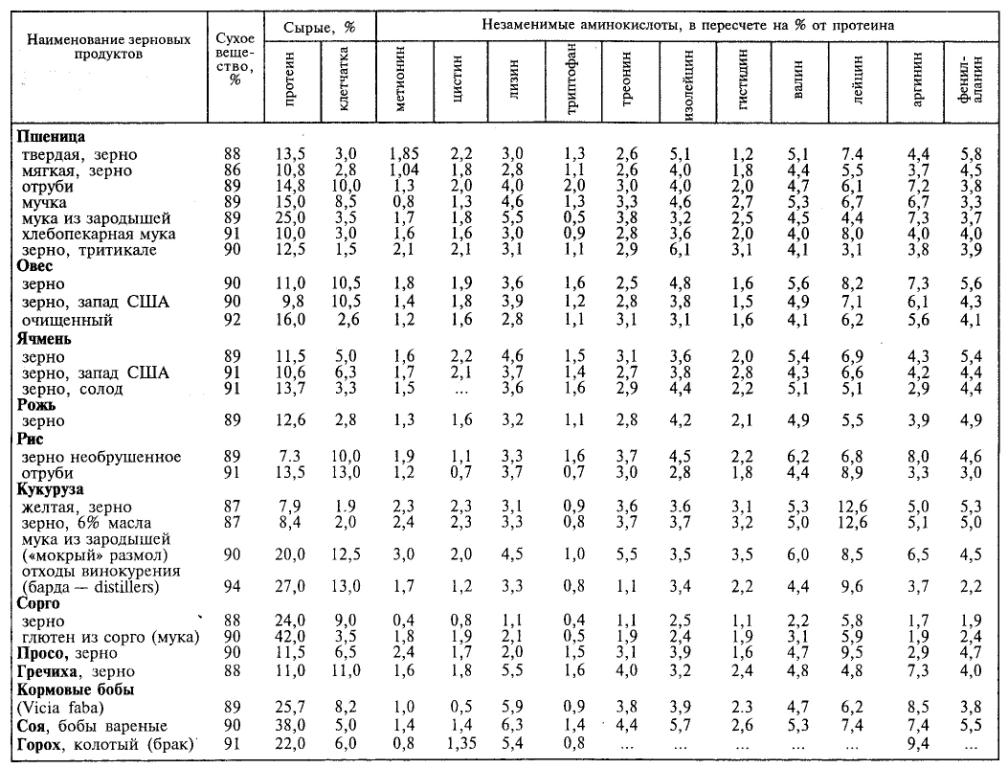
Аминокислотный состав зерна озимого тритикале и озимой ржи характеризуется значительным содержанием пролина (0,94 и 0,79 г/100 г) и лейцина (1,62 и 1,52 г/100 г). Устойчивость к засухе и другим неблагоприятным факторам многие исследователи связывают с содержанием пролина в тканях растений, который активно синтезируется в ответ на разные стрессовые воздействия, выступая в качестве осмопро-тектора [13-17]. Учитывая, что с увеличением накопления пролина растения озимых культур откликаются на воздействия различных неблагоприятных факторов, можно сделать вывод о том, что сорта озимого тритикале и озимой ржи сильнее лимитируются условиями произрастания, чем озимая пшеница. Массовая доля пролина в зерне сортов озимой пшеницы в среднем составляет 0,32 г/100 г, что на 0,62 и 0,47% меньше, чем у озимого тритикале и озимой ржи соответственно.
Суммарное содержание незаменимых аминокислот в озимой ржи меньше на 16,9%, чем в зерне озимого тритикале, и на 5%, чем в зерне озимой пшеницы (табл. 3). Белки злаковых культур неполноценны по ряду незаменимых аминокислот, прежде всего по лизину, метионину, триптофану и треонину. Поэтому при оценке аминокислотной сбалансированности используют эти четыре аминокислоты.
3). Белки злаковых культур неполноценны по ряду незаменимых аминокислот, прежде всего по лизину, метионину, триптофану и треонину. Поэтому при оценке аминокислотной сбалансированности используют эти четыре аминокислоты.
По такой лимитирующей незаменимой аминокислоте, как треонин, зерно озимой пшеницы богаче по сравнению с озимой рожью и озимым тритикале на 18 и 12%, по метионину — на 64,0 и 22,6%, по лизину — на 44 и 14% соответственно. Триптофана накапливается в зерне мало — 0,11-013 г/100 г.
Таким образом, в зерне озимых злаков, произрастающих на территории лесостепи юго-востока Западной Сибири, содержание лимитирующих незаменимых аминокислот у озимой пшеницы больше, чем у остальных изучаемых культур. Наиболее дефицитной по содержанию в белке треонина, метионина и лизина является озимая рожь. У белков всех изучаемых культур первой лимитирующей незаменимой аминокислотой является метионин, второй — лизин.
По суммарному содержанию незаменимых аминокислот белок озимого тритикале несколько отличается от озимой пшеницы, сильнее — от озимой ржи.
Расчет аминокислотного скора показал, что в наибольшей степени требованиям ФАО/ВОЗ соответствуют сорта озимой ржи Петровна 1 и гибрида (Р1) Хеллтоп. У этих культур уровень качества зерна приближается к высокому. Биологическая ценность зерна составляет 17 и 18% соответственно (табл. 4).
Биологическую ценность белка лимитирует незаменимая аминокислота с наименьшим аминокислотным скором. У всех изучаемых сортов озимых культур такой аминокислотой является лизин, аминокислотный скор которой составляет менее 100%. У сортов озимой пшеницы Новосибирская 2 и Новосибирская 3 содержание лизина высокое в сравнении с другими озимыми культурами — аминокислотный скор составляет 84 и 80% соответственно. Отмечается низкий аминокислотный скор у сортов озимой ржи, особенно у сорта Петровна 1 — всего 32%.
Результаты определения аминокислотного скора в исследуемых образцах зерна озимых культур показали, что по биологической ценности зерно сортов, произрастающих в условиях лесостепи юго-востока Западной Сибири, имеет средний и низкий уровни качества.
Таблица 3. Содержание незаменимых аминокислот в зерне озимых культур (пшеницы, ржи и тритикале),
г /100 г
Незаменимые аминокислоты Суточная потребность, г Озимая пшеница Озимая рожь Озимого тритикале
содержание, г/100 г % от дневной нормы содержание, г/100 г % от дневной нормы содержание, г/100 г % от дневной нормы
Треонин 2,5 0,77 19,3 0,63 15,8 0,68 17,0
Валин 3,5 0,68 13,6 0,61 12,2 0,61 12,2
Метионин 3 0,31 8,9 0,11 ЗД 0,24 6,9
Изолейцин 3,5 0,65 16,3 0,39 9,8 0,56 14,0
Лейцин 5 0,55 7,9 1,52 21,7 1,62 23,1
Триптофан 1 0,11 11,0 0,13 13,0 0,13 13,0
Фенил ал анин 3 0,68 13,6 0,34 6,8 0,57 11,4
Лизин 4 0,43 7,8 0,24 4,4 0,37 6,7
Таблица 4. Аминокислотный скор озимой пшеницы, озимой ржи и озимого тритикале, %
Аминокислотный скор озимой пшеницы, озимой ржи и озимого тритикале, %
Незаменимая аминокислота Сорта озимой пшеницы Гибрид и сорта озимой ржи Сорта озимого тритикале
Скипетр Новосибирская 2 Новосибирская 3 Зу Драйве Хеллтоп (Fi) Петровна 1 Алтайская 5 Омская
Валил 118 139 130 128 108 128 107 120
Изолейцин 152 175 165 97 97 90 135 133
Лейцин 230 207 227 167 264 220 247 216
Лизин 73 84 80 56 44 32 76 56
Метионин + цистин 131 162 174 86 100 94 142 103
Треонин 160 215 203 175 140 160 165 177
Триптофан 120 90 110 130 150 110 120 140
Фенилаланин + 150 161 170 142 45 83 180 88
тирозин
Биологическая цен- 31 30 23 33 18 17 29 26
ность белка (КРАС)
Установлено, что самым низким уровнем качества зерна обладает сорт озимой ржи Зу Драйве иностранной селекции. Биологическая ценность его составляет 33%, что по градации [11] соответствует низкому уровню качества. Зерно сортов низкого качества используют преимущественно на фуражные цели. Например, в Канаде, США и других странах такие сорта не допускаются к районированию [18].
Биологическая ценность его составляет 33%, что по градации [11] соответствует низкому уровню качества. Зерно сортов низкого качества используют преимущественно на фуражные цели. Например, в Канаде, США и других странах такие сорта не допускаются к районированию [18].
Выводы
В результате проведенных исследований установлено, что качество зерна озимой пшеницы, озимой ржи и озимого тритикале, произрастающих на территории лесостепи юго-востока Западной Сибири, далеко от максимально возможного. Анализ содержания белка и аминокислот показал биосинтетические особенности формирования зерна изучаемых видов и сортов озимых культур.
Зерно изучаемых культур характеризуется различным содержанием белка и его аминокислотным составом. Сравнение качества зерна сортов озимых злаковых культур позволило установить, что сорт озимой пшеницы Новосибирская 3 характеризуется низким содержанием белка (11,3%), но является наиболее биологически ценным по сравнению с другими изучаемыми сортами — Новосибирская 2 и Скипетр. Низким содержанием белка отличаются сорт озимой ржи иностранной селекции Зу Драйве (10,14%) и гибрид (Fi) Хеллтоп (10,34%). Однако белок зерна гибрида Хеллтоп имеет более высокую биологическую ценность по сравнению с сортом Зу Драйве. Высокобелковыми оказались сорта озимого тритикале сибирской селекции. В среднем они накапливают белка до 14,2% и формируют зерно среднего уровня качества. Исходя из вышеизложенного, следует отметить, что изученные сорта озимых злаковых культур, произрастающие на территории лесостепи юго-востока Западной Сибири, не обеспечивают потребность региона в накоплении полноценного белка, а значит, и биологически полноценного питания. Полученные результаты позволяют ближе подойти к пониманию причин получения низкого и среднего качества зерна на исследуемой территории, как к явлению, широко распространенному во многих регионах Российской Федерации, и указывают на новые возможности нормализации метаболических процессов формирующейся зерновки. Учитывая особую значимость зерна как источника белка и незаменимых аминокислот для человека и животных, следует проводить исследования с целью выявления сортов озимых злаковых культур с максимальным содержанием биологически важных веществ.
Низким содержанием белка отличаются сорт озимой ржи иностранной селекции Зу Драйве (10,14%) и гибрид (Fi) Хеллтоп (10,34%). Однако белок зерна гибрида Хеллтоп имеет более высокую биологическую ценность по сравнению с сортом Зу Драйве. Высокобелковыми оказались сорта озимого тритикале сибирской селекции. В среднем они накапливают белка до 14,2% и формируют зерно среднего уровня качества. Исходя из вышеизложенного, следует отметить, что изученные сорта озимых злаковых культур, произрастающие на территории лесостепи юго-востока Западной Сибири, не обеспечивают потребность региона в накоплении полноценного белка, а значит, и биологически полноценного питания. Полученные результаты позволяют ближе подойти к пониманию причин получения низкого и среднего качества зерна на исследуемой территории, как к явлению, широко распространенному во многих регионах Российской Федерации, и указывают на новые возможности нормализации метаболических процессов формирующейся зерновки. Учитывая особую значимость зерна как источника белка и незаменимых аминокислот для человека и животных, следует проводить исследования с целью выявления сортов озимых злаковых культур с максимальным содержанием биологически важных веществ.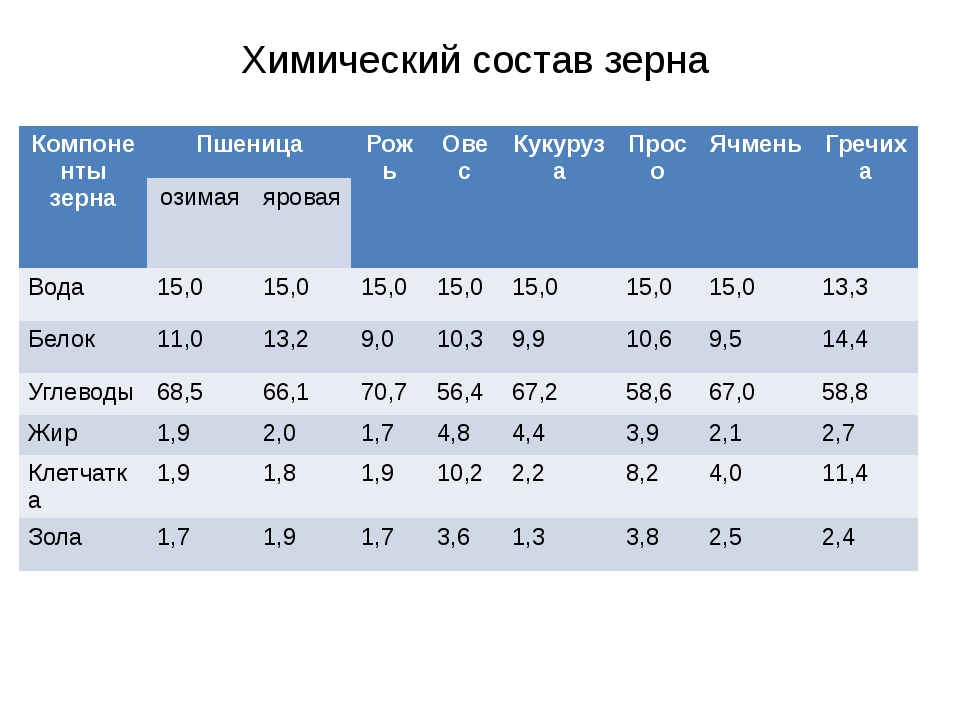 17.
17.
3. Корякин В.В., Солодова Ю.П. Динамика посевных площадей озимой ржи и значимость как злаковой культуры в решении продовольственной программы // Вестник ТГУ. 2011. Т. 16, вып. 2. С. 660-662.
4. Белугина Н.О., Блинова Е.В. Источники хозяйственно-ценных признаков озимой ржи // Труды по прикладной ботанике, генетике и селекции. 2006. Т. 162. С. 166-170.
5. Минеев A.B., Дебрецени Б., Мазур Т. Биологическое земледелие и минеральные удобрения. М., 1993. 415 с.
6. Исмагилов P.P., Ахиярова Л.М., Гайсина Л.Ф. Содержание пентозанов в зерне гибридов озимой ржи // Вави-ловские чтения : материалы Межд. науч.-практ. конф. Саратов, 2012. С. 89-91.
7. Силин М.А. Ценный кормовой продукт из зерна ржи // Кормопроизводство. 2012. №3. С. 44.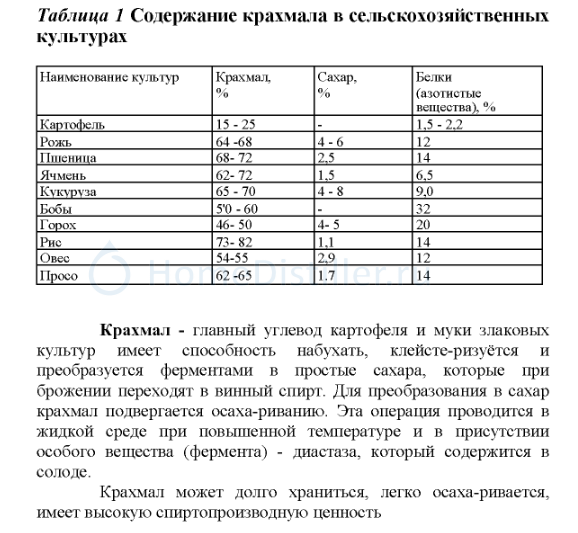
8. Босиева О.И., Плиева Е.А., Джиоева Г.Ф. Содержание белка и аминокислотный состав зерна тритикале // Известия Горского ГАУ. 2011. Т. 48, №2. С. 102-104.
9. Мединский А.В., Степочкин П.И. Изучение озимых тритикале в Сибирском научно-исследовательском институте растениеводства и селекции // Сибирский вестник сельскохозяйственной науки. 2014. №1. С. 32-35.
10. Рядчиков В.Г. Улучшение зерновых белков и их оценка. М., 1998. 368 с.
11. Патент 2198538 (РФ). Способ оценки биологической ценности растительного белка / А.П. Стаценко. БИ. 2001. №8. С. 86.
12. Хелдт Г.В. Биохимия растений : пер. с англ. М., 2011. 471 с.
13. Демин Д.А. Влияние поражения твердой головней на количество и фракционный состав белков в зерне пшеницы // Актуальные проблемы сельскохозяйственной науки и образования : сб. науч. тр. Самара, 2005. Вып. 4. С. 154-157.
науч. тр. Самара, 2005. Вып. 4. С. 154-157.
14. Gutterman Y. Survival strategies of annual desert plants : Adaptations of desert organisms. Berlin ; New York, 2002. Pp. 1-36.
15. Verbruggen N., Hermans C. Proline accumulation in plants : A. Review // Amino acids. 2008. Vol. 35. Pp. 753-759.
16. Szabados L., Savoure A. Proline : multifunctionae amino acid // Trends Plant Sci. 2010. Vol. 15. Pp. 89-97.
17. Lehmann S., Funck D., Szabados L., Rentsch D. Proline Metabolism and Transport in Plant Development // Amino acids. 2010. Vol. 39. Pp. 949-962.
18. Брежнев Д.Д., Шмараев Г.Е. Селекция растений в США. М., 1976. 349 с.
Поступило в редакцию 21 мая 2015 г. После переработки 20 сентября 2015 г.
После переработки 20 сентября 2015 г.
Kondratenko E.P.1, Konstantinova O.B.1, Soboleva O.M.1 , Izhmulkina E.A.1, Verbitskaya N.V.1, Sukhikh A.S.2 THE CONTENT OF PROTEIN AND AMINO ACIDS IN GRAIN OF WINTER CROPS GROWING ON THE TERRITORY OF FOREST-STEPPE SOUTH-EAST OF WESTERN SIBERIA
Kemerovo state agricultural Institute, st. Markovtseva, 5, Kemerovo, 650056 (Russia), e-mail: [email protected] 2Kemerovo state medical Academy, st. Voroshilova, 224, Kemerovo, 650029 (Russia)
This article presents the results of the analysis of the content of protein and amino acids in grain of winter wheat, winter rye and winter triticale growing on the territory of forest-steppe South-East of Western Siberia depending on the type of culture and biological characteristics of the variety. The qualitative and quantitative amino acid composition of three grain species and 8 varieties of winter cereals.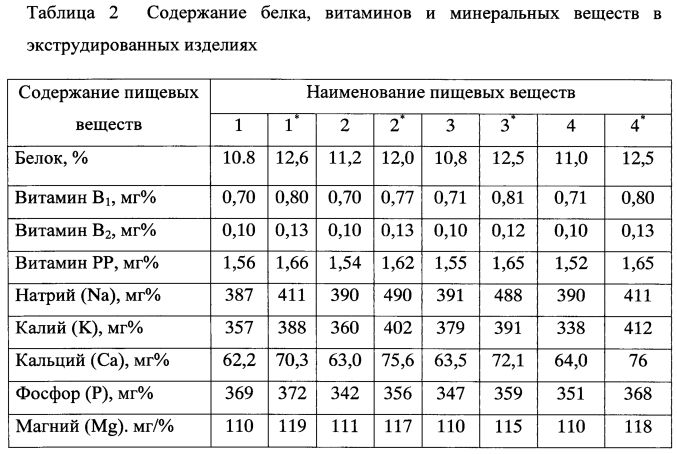 Hilltop respectively. High protein was characterized by varieties of winter triticale of the Altai 5 — 14,31% and Omsk 14,08%). The content of all amino acids in the grain of winter rye grain is inferior to the other studied cultures the necessity of the required analysis of the amino acid composition of grain food crops to determine their biological value.
Hilltop respectively. High protein was characterized by varieties of winter triticale of the Altai 5 — 14,31% and Omsk 14,08%). The content of all amino acids in the grain of winter rye grain is inferior to the other studied cultures the necessity of the required analysis of the amino acid composition of grain food crops to determine their biological value.
Keywords: grade, grain, winter wheat, winter rye, winter triticale, protein, amino acids, near the amino acid, biological value.
References
1. Vysotskii V.G., IatsyshinaT.A., Rymarenko T.Y. Meditsinskii referativnyi zhumal, 1976, no. 6, pp. 24-35. (inRuss.).
2. Volkov O.V. Sovremennye problemy tekhnologii proizvodstva, khraneniia, pererabotki i ekspertizy sel’sko-khoziaistvennoi produktsii : materialy mezhdunar. nauch.-prakt.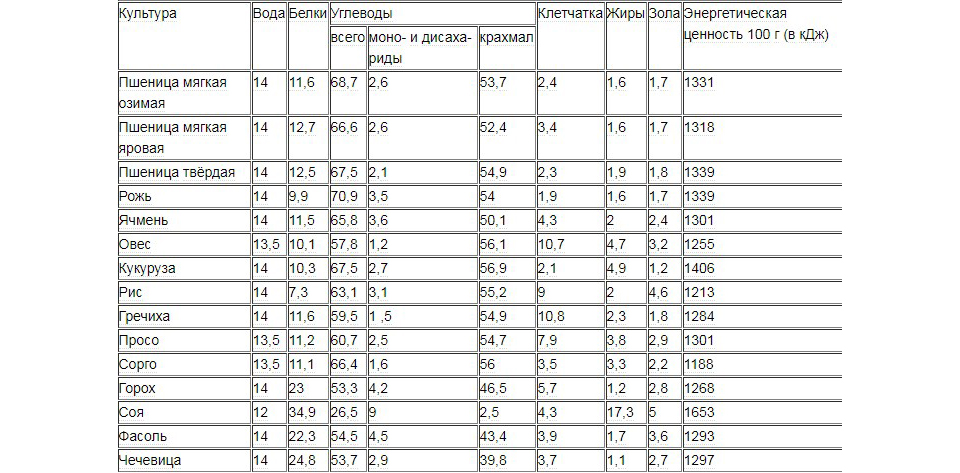 7. (inRuss.).
7. (inRuss.).
3. Koriakin V. V., Solodova Iu.P. Vestnik TGU, 2011, vol. 16, no. 2, pp. 660-662. (in Russ.).
4. Belugina N.O., Blinova E. V. Trudypoprikladnoi botanike, genetike i selektsii, 2006, vol. 162, pp. 166-170. (in Russ.).
5. Mineev A.V., Debretseni B., Mazur T. Biologicheskoe zemledelie i mineral’nye udobreniia. [Biological agriculture and fertilizers], Moscow, 1993, 415 p. (inRuss.).
6. Ismagilov R.R., Akhiiarova L.M., Gaisina L.F. Vavilovskie chteniia : materialy mezhd. nauch.-prakt. konf. [Vavilov Reading : Materials of the international scientific-practical conference], Saratov, 2012, pp. 89-91. (inRuss.).
7. Silin M.A. Kormoproizvodstvo, 2012, no. 3, pp. 44. (in Russ.).
8. Bosieva O.I., Plieva E.A., Dzhioeva G.F. Izvestiia Gorskogo GAU, 2011, vol. 48, no. 2, pp. 102-104. (inRuss.).
Bosieva O.I., Plieva E.A., Dzhioeva G.F. Izvestiia Gorskogo GAU, 2011, vol. 48, no. 2, pp. 102-104. (inRuss.).
9. Medinskii A.V., Stepochkin P.I. Sibirskii vestniksel’skokhoziaistvennoi nauki, 2014, no. 1, pp. 32-35. (inRuss.).
10. Riadchikov V.G. Uluchshenie zernovykh belkov i ikh otsenka. [Improving cereal proteins and their evaluation], Moscow, 1998, 368p. (inRuss.).
11. Patent 2198538 (RU). 2001. (in Russ.).
12. Kheldt G.V. Biokhimiia rastenii. [Plant Biochemistry], Moscow, 2011, 471 p. (inRuss.).
13. Demin D.A. Aktual’nye problemy sel’skokhoziaistvennoi nauki i obrazovaniia : sb. nauch. tr. [Actual problems of agricultural science and education: collection of scientific papers]. Samara, 2005, no. 4, pp. 154-157. (inRuss. ).
).
14. Gutterman Y. Survival strategies of annual desert plants : Adaptations of desert organisms. Berlin ; New York, 2002, pp. 1-36.
15. Verbruggen N., Hermans C. Amino acids, 2008, vol. 35, pp. 753-759.
16. Szabados L., Savoure A. Trends Plant Sci., 2010, vol. 15, pp. 89-97.
17. Lehmann S., Funck D., Szabados L., Rentsch D. Amino acids, 2010, vol. 39, pp. 949-962.
18. Brezhnev D.D., Shmaraev G.E. Selektsiia rastenii v SShA. [Plant breeding in the United States], Moscow, 1976, 349p. (inRuss.).
Received May 21, 2015 Revised September 20, 2015
Corresponding author.
Зерновые корма и продукты их переработки
Зерновые корма и продукты их переработки служат основными источниками энергии, белковых веществ и крахмала. В определенные периоды роста свиней, например при доращивании и откорме, эти корма могут составлять около 90% питательности рациона. Однако, несмотря на возможность включения зерна злаковых в рационы в большом количестве, некоторые особенности его химического состава и питательности требуют тщательного балансирования рационов в соответствии с потребностями свиней, используя различные дополнители.
В определенные периоды роста свиней, например при доращивании и откорме, эти корма могут составлять около 90% питательности рациона. Однако, несмотря на возможность включения зерна злаковых в рационы в большом количестве, некоторые особенности его химического состава и питательности требуют тщательного балансирования рационов в соответствии с потребностями свиней, используя различные дополнители.
В зерне злаковых сухого вещества содержится 82—90%, количество его зависит от многих факторов, в первую очередь от способа уборки и хранения.
Содержание протеина в зерне может колебаться от 5 до 26%, что обусловливается сортовыми особенностями растений, климатическими факторами, условиями выращивания, сроками и способами уборки и т. д. Невысокий в среднем уровень протеина в зерне злаковых является одним из факторов, ограничивающих применение его в качестве единственного корма в рационах свиней. К тому же протеин зерна злаков имеет низкую биологическую ценность, которая составляет для пшеницы 62—67%, овса — 68, ячменя — 64, проса — 57, кукурузы — 52—58, риса — 83—86% (по Н. Н. Иванову). Установлено, что ценность белка хлебных злаков для стимуляции роста располагается в следующем убывающем порядке: овес, ячмень, кукуруза или пшеница.
Н. Иванову). Установлено, что ценность белка хлебных злаков для стимуляции роста располагается в следующем убывающем порядке: овес, ячмень, кукуруза или пшеница.
Количество углеводов в этом корме, которые в основном представлены крахмалом, сахарами, клетчаткой, гемицеллюлозами, пентозанами, достигает 80% сухого вещества. Крахмал находится лишь в мучнистой части эндосперма, а в пленках, оболочках, алейроновом слое и зародышах его или нет, или очень мало. Содержание крахмала изменяется в зерне пшеницы в пределах 49—73%, ржи — 55—73, ячменя 45—68,-овса — 24—64, кукурузы — 61—83, проса — 51—70, риса — 48—68,-а в полированном рисе — 71—86%. Количество крахмала в зернах одной и той же культуры и сорта выше в северных влажных районах, чем в южных. Сахар в основном находится в зародышах семян в довольно небольшом количестве — 2—5%, но если в овсе он составляет 0,6—2,2%, то в семенах ржи может достигать 9%. Клетчатка — основной компонент семенных оболочек и клеточных стенок. Содержание ее зависит главным образом от пленчатости зерна; например, в пленчатом яровом овсе она составляет от 7 до 24%, а в голозерном овсе — в среднем 4,7%. В недозрелом зерне любого вида процент клетчатки повышен.
В недозрелом зерне любого вида процент клетчатки повышен.
На долю жира в зерне пшеницы, ржи, ячменя и риса приходится 1,6—3,2%, овса, кукурузы — 3—8, проса —3,0—5,2%. Богаты им зародыши семян. Группа липоидов представлена стеринами, лецитинами, кефалином.
Минеральный состав семян зерновых злаковых в большой степени зависит от содержания и формы минеральных элементов в почве. Фосфора в зерне 0,27—0,39, но значительная часть его находится в виде фитина, обладающего способностью связывать пищевой кальций, а возможно, и магний. Зерно злаковых служит важным источником калия и магния, сера входит в основном в состав серосодержащих аминокислот — цистина, цистеина, метионина. Кроме названных элементов, в золе зерна злаковых обнаруживается марганец, медь, цинк, бор, алюминий, йод, кобальт, никель, молибден, фтор, селен, бром, титан, олово, мышьяк, литий, ванадий, барий, стронций, цезий, рубидий и другие, многие из которых являются биогенными элементами.
Каротином довольно богаты желтозерная кукуруза и продукты ее переработки.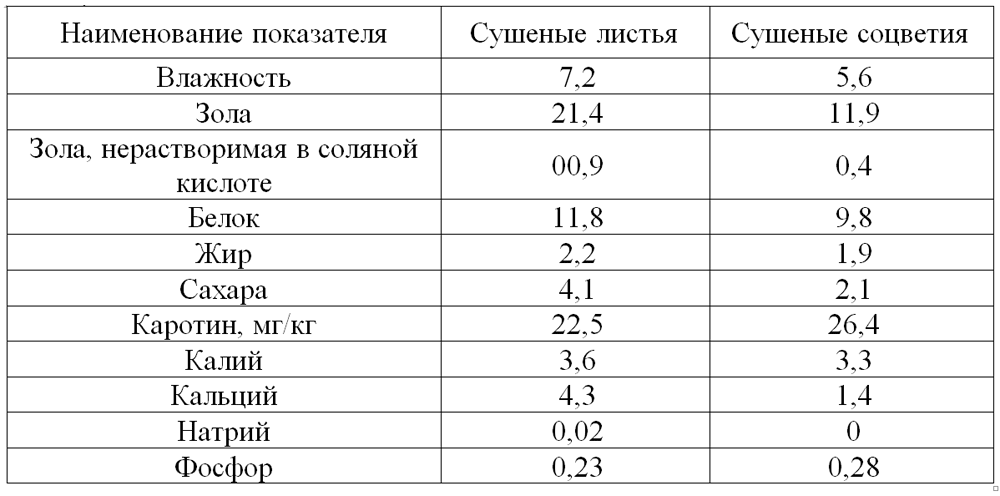 В других культурах он или отсутствует, или его содержится очень мало (ячмень). В 1 кг зерна злаковых в среднем около 10 мг витамина Е, который находится в основном в зародышах. По содержанию его только семена некоторых бобовых культур и зеленые листья растений превосходят зерно злаковых. Витамин D практически отсутствует. Зерно злаковых представляет собой хороший источник витамина В1, уступая лишь семенам некоторых зернобобовых культур и дрожжам. Витамином В1 богаты зародыши зерна и алейроновый слой, в мучнистой части эндосперма его значительно меньше. Витамином В2 оно бедно. Витамина В6 в зерне злаков в среднем около 5 мг. Особенно много его в рисовых отрубях — 300—500 мг. Витамин РР (никотиновая кислота) имеется в зерне всех злаков в довольно значительном количестве. Содержание никотиновой кислоты обычно больше в отрубях злаковых, чем в эндосперме. Пантотеновой кислоты в эндосперме зерна обычно в 2—4 раза меньше, чем в зародышах и алейроновом слое. Химический состав зерновых злаковых и их питательность приведены в таблице
В других культурах он или отсутствует, или его содержится очень мало (ячмень). В 1 кг зерна злаковых в среднем около 10 мг витамина Е, который находится в основном в зародышах. По содержанию его только семена некоторых бобовых культур и зеленые листья растений превосходят зерно злаковых. Витамин D практически отсутствует. Зерно злаковых представляет собой хороший источник витамина В1, уступая лишь семенам некоторых зернобобовых культур и дрожжам. Витамином В1 богаты зародыши зерна и алейроновый слой, в мучнистой части эндосперма его значительно меньше. Витамином В2 оно бедно. Витамина В6 в зерне злаков в среднем около 5 мг. Особенно много его в рисовых отрубях — 300—500 мг. Витамин РР (никотиновая кислота) имеется в зерне всех злаков в довольно значительном количестве. Содержание никотиновой кислоты обычно больше в отрубях злаковых, чем в эндосперме. Пантотеновой кислоты в эндосперме зерна обычно в 2—4 раза меньше, чем в зародышах и алейроновом слое. Химический состав зерновых злаковых и их питательность приведены в таблице
Повышение содержания и качества белка в зерне
На оба из компонентов влияет уровень запасов азота и серы в тканях растения на разных этапах развития. Известно, что при высоких показателях урожайности содержание белка в зернах понижено, поэтому очень важно заранее оценить прогнозируемую урожайность, еще на этапе налива зерна. При управлении качеством белка основной задачей является выработка растением высокомолекулярного белка глютена. Благодаря компонентам глютена, например, таким как глиадин, глютенин, альбумин и глобулин, изделия из пшеничной муки имеют уникальную расширяемость и высокие технологические свойства.
Известно, что при высоких показателях урожайности содержание белка в зернах понижено, поэтому очень важно заранее оценить прогнозируемую урожайность, еще на этапе налива зерна. При управлении качеством белка основной задачей является выработка растением высокомолекулярного белка глютена. Благодаря компонентам глютена, например, таким как глиадин, глютенин, альбумин и глобулин, изделия из пшеничной муки имеют уникальную расширяемость и высокие технологические свойства.
С помощью агрономических технологий производитель должен обеспечить оптимальное внесение азотных удобрений в необходимых нормах и в срок, когда растения еще способны доставить его в зерна. Кроме того, необходимо обеспечить соответствующее серное питания, что не ограничивает эффективность использования азота.
С помощью агрономических технологий производитель должен обеспечить оптимальное внесение азотных удобрений в необходимой дозе и оптимальные сроки, когда растения еще способны доставить его в зерна. Кроме того, необходимо обеспечить соответствующее серное питание, не ограничивающее эффективность использования азота.
Кроме того, необходимо обеспечить соответствующее серное питание, не ограничивающее эффективность использования азота.
Азот является основным компонентом аминокислот – составных элементов белка в зерне пшеницы. Таким образом, поддержание его запасов имеет решающее значение при выращивании пшеницы с высокими показателями содержания белка. По содержанию белка в зерне можно определить, получают посевы оптимальное азотное питание. При изменении норм внесения азота (из-за экономических трудностей), или если вносимый азот отличается от оптимального, уровень содержания белка в зернах обычно изменяется примерно на 1% на каждые 50 кг N/га.
Для получения более высокого содержания белка потребуется увеличение вносимых доз азота в связи с более высокими потребностями. Общая потребность растений в азоте зависит от назначения использования зерна, так же как и содержание в нем белка.
Содержание белка в зернах кормовой пшеницы при оптимальном азотном питании обычно составляет около 11% (что эквивалентно 1,9% N). Однако хлебные сорта пшеницы дают оптимальные результаты урожая с содержанием белка около 12%, поэтому для достижения рыночного показателя выше 13% часто требуется дополнительное внесение азотных удобрений. Пониженное содержание белка, то есть менее 10% для кормовых сортов пшеницы, указывает на субоптимальне использования азота.
Однако хлебные сорта пшеницы дают оптимальные результаты урожая с содержанием белка около 12%, поэтому для достижения рыночного показателя выше 13% часто требуется дополнительное внесение азотных удобрений. Пониженное содержание белка, то есть менее 10% для кормовых сортов пшеницы, указывает на субоптимальне использования азота.
Важным фактором является срок внесения азотных удобрений. При раннем внесении азота (фазы 25-32 по Задоксу) производитель получит более высокую урожайность, в то время,как для повышения содержания белка внесение азотных удобрений должно быть осуществлено между 37 и 59 фазой по шкале Задокса.
При недостатке запасов азота в тканях листьев могут применяться лечебные мероприятия в виде азотного внекорневой подкормки, спланированного по времени к фазе цветения (фаза 70 по Задоксу).
Сера является одним из основных структурных элементов белков и, следовательно, от этого элемента во многом зависит уровень содержания и качество белков зерна. Сера также входит в состав нескольких основных аминокислот, которые обеспечивают ценность пшеничной муки. Такими аминокислотами являются цистеин, метионин, треонин и лизин.
Сера также входит в состав нескольких основных аминокислот, которые обеспечивают ценность пшеничной муки. Такими аминокислотами являются цистеин, метионин, треонин и лизин.
Марганец и цинк играют важную роль в процессах метаболизма растений, включая азотный обмен. Улучшение азотного обмена способствует увеличению количества азота, участвует в синтезе белков.
Пищевая ценность зерновых и масличных культур
Значение пищевых продуктов в питании человека определяется их химическим составом. Зерно каждой культуры в зависимости от сорта, условий выращивания и других факторов может иметь различный химический состав, но при любых условиях в зерне имеются практически все необходимые вещества для развития человеческого организма.
Такие зерновые культуры, как пшеница, рожь, гречиха, ячмень, просо, кукуруза и соя широко распространены на территории РФ и, кроме того, за счет достаточно высокого содержания углеводов, белков, жиров, минеральных веществ и витаминов являются ценным и недорогим сырьем для производства пищевых продуктов.
Биохимический состав зерна характеризуется макронутриентами — углеводами, белками, жирами и микронутриентами — минеральными веществами, витаминами и другими биологически активными компонентами. В первую очередь химический состав зерна определяется видом, однако он может существенно меняться в зависимости от места и условий выращивания. Средний химический состав зернобобовых и масличных культур приведен в табл. 1.
Таблица 1. Средний химический состав различных культур (%)
| Культуры | Вода | Белки | Жиры | Углеводы | Зола | Энергетическая ценность, ккал | ||
| Моно- и дисахариды | Крахмал | Клетчатка | ||||||
| Злаковые | ||||||||
| Пшеница: | ||||||||
| мягкая озимая | 14,0 | 11,6 | 2,1 | 1,2 | 53,7 | 2,6 | 1,7 | 290 |
| мягкая яровая | 14,0 | 12,5 | 2,3 | 0,9 | 53,0 | 2,5 | 1,7 | 291 |
| твердая | 14,0 | 13,0 | 2,5 | 0,8 | 54,5 | 2,3 | 1,7 | 301 |
| Рожь | 14,0 | 9,9 | 2,5 | 1,5 | 54 | 2,6 | 1,7 | 287 |
| Тритикале | 14,0 | 12,8 | 2,1 | 1,0 | 53,5 | 2,6 | 1,7 | 293 |
| Овес | 13,5 | 10,0 | 6,2 | 1,1 | 36,5 | 107 | 3,2 | 250 |
| Ячмень | 14,0 | 10,3 | 2,4 | 1,3 | 48,1 | 4,3 | 2,4 | 264 |
| Просо | 13,5 | 11,2 | 3,9 | 1,9 | 54,7 | 7,9 | 2,9 | 311 |
| Гречиха | 14,0 | 11,6 | 3,2 | 1,5 | 54,9 | 10,8 | 1,8 | 295 |
| Рис | 14,0 | 7,4 | 2,6 | 0,9 | 55,2 | 9,0 | 3,9 | 283 |
| Сорго | 13,5 | 10,6 | 4,1 | 1,6 | 58,0 | 3,5 | 2,2 | 323 |
| Кукуруза: | ||||||||
| зубовидная | 14,0 | 8,3 | 4,0 | 1,6 | 59,8 | 2,1 | 1,2 | 320 |
| кремнистая | 14,0 | 9,2 | 4,2 | 1,6 | 57,3 | 2,2 | 1,2 | 316 |
| сахарная | 14,0 | 11,2 | 4,5 | 8,0 | 29,9 | 2,5 | 1,3 | 338 |
| лопающаяся | 14,0 | 10,7 | 4,3 | 3,0 | 55 | 2,0 | 1,1 | 318 |
| Амарант | 11,3 | 13,6 | 7,0 | 58,5 | — | 2,9 | 371 | |
| Бобовые | ||||||||
| Горох | 14,0 | 23 | 2,0 | 4,6 | 46,5 | 5,7 | 2,8 | 298 |
| Фасоль | 14,0 | 22,3 | 2,0 | 3,2 | 43,4 | 3,9 | 3,6 | 292 |
| Чечевица | 14,0 | 24,8 | 1,5 | 2,9 | 39,8 | 3,7 | 2,7 | 284 |
| Соя | 12,0 | 34,9 | 17,3 | 5,7 | 3,5 | 4,3 | 5 | 332 |
| Нут | 14,0 | 20,1 | 5,0 | 6,2 | 43,2 | 3,7 | 3 | 329 |
| Масличные | ||||||||
| Подсолнечник | 11,0 | 14,8 | 40,8 | 16 | 14,5 | 2,9 | 601 | |
| Лен | 8,0 | 24,1 | 48,6 | 11,1 | 2,4 | 3,8 | — | |
| Кунжут | — | 19,4 | 48,7 | 12,2 | 2,9 | 5,1 | — | |
| Клещевина | 7,0 | 18,3 | 51,3 | 2,0 | 18,6 | 2,8 | — | |
| Кориандр | 10,0 | 14,8 | 22,1 | 28,0 | 18,6 | 5,6 | — | |
Углеводы составляют главную массу зерна и семян — примерно две трети. Это основной питательный и опорный материал растительных клеток и тканей. Велика их роль в питании человека; они представляют собой энергетический материал — главный источник калорий. Важнейшие углеводы, встречающиеся в зерне, представлены на рис. 1.
Это основной питательный и опорный материал растительных клеток и тканей. Велика их роль в питании человека; они представляют собой энергетический материал — главный источник калорий. Важнейшие углеводы, встречающиеся в зерне, представлены на рис. 1.
Рис. 1. Классификация углеводов зерна
По пищевой ценности углеводы делятся на усвояемые и неусвояемые. К первым относятся глюкоза, фруктоза, сахароза, лактоза, мальтоза и альфа-глюкановые полисахариды — крахмал, декстрины и гликоген. Ко вторым — целлюлоза, гемицеллюлоза, пектиновые вещества, камеди и слизи.
В последнее время повышенное внимание привлекают полисахариды второго порядка — целлюлоза (клетчатка), гемицеллюлоза (полуклетчатка) и пектин. Вместе с лигнином они объединяются в группу веществ, названных пищевыми волокнами, которые плохо усваиваются организмом человека, из-за чего их часто называют балластными веществами. При этом они играют важную роль в процессах функционирования пищеварительного тракта.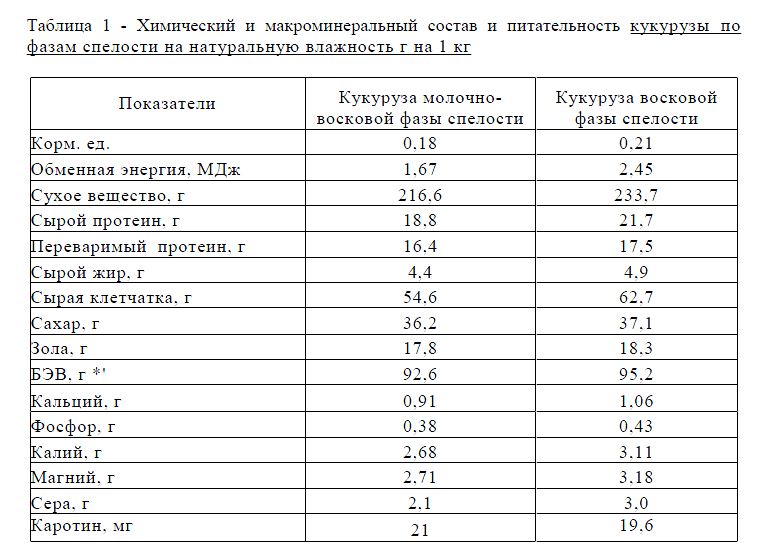
Пищевые волокна являются строительным материалом стенок клетки растений. Клетчатка — наиболее распространенный углевод с высокой молекулярной массой — образует структурную основу оболочек растительных клеток. Основное ее физиологическое назначение — связывать воду. В зависимости от происхождения и степени предварительной обработки усвояемость клетчатки составляет от 6 до 23 %, перевариваемость клетчатки пшеничных отрубей около 15 %.
В стенках клетки зерна каркас из клетчатки заполнен гемицеллюлозой и лигнином. Гемицеллюлоза по своему функциональному назначению и свойствам занимает промежуточное положение между клетчаткой и крахмалом. С одной стороны, как и клетчатка, она является строительным материалом стенок клетки, с другой — запасным питательным веществом, как и крахмал. Гемицеллюлоза нерастворима в воде, способна удерживать воду и связывать катионы, ее перевариваемость составляет 69…95 %. Среднее содержание клетчатки и гемицеллюлозы в зерне представлено в табл. 2.
Таблица 2. Содержание полисахаридов в зернобобовых культурах (% на сухое вещество)
| Культуры | Клетчатка | Гемицеллюлоза (пентозаны) | Пектиновые вещества |
| Пшеница | 2,0…3,4 | 5,0…8,0 | 0,5 |
| Рожь | 1,9…2,8 | 9,0…11,0 | 0,6 |
| Овес (в пленках) | 10,5…16,5 | 12,0…14,0 | 1,0 |
| Ячмень (в пленках) | 4,0…6,0 | 9,0…12,0 | 0,4 |
| Горох | 4,5…6,5 | 4,4…8,0 | 3,0 |
| Фасоль | 3,8 | 3,9 | 3,7 |
| Соя | 4,3 | 6,3 | — |
Пектиновые вещества, или пектины, содержатся в первичных клеточных стенках и межклеточных тканях. Водонерастворимая форма пектина называется протопектином. Одно из важнейших свойств пектина — способность образовывать комплексы при взаимодействии с тяжелыми металлами и радионуклидами. Это особенно важно для питания лиц, имеющих контакт с неблагоприятной радиационной средой или средой, загрязненной тяжелыми металлами.
Лигнин — высокомолекулярное соединение, состоящее из полимеров ароматических спиртов. В организме это вещество способно связывать соли желчных кислот и другие органические вещества.
Белки — сложные высокомолекулярные органические вещества со строго определенным составом и строением. Они содержат углерод, азот, водород и серу. С белками связаны все основные жизненные процессы организма. Они участвуют в образовании иммунных тел, передаче генетической информации, транспортировании веществ в организме, регулируют и катализируют биохимические реакции в процессе обмена веществ. Около 40 % белковой массы продукта используется как источник энергии, а остальное — как пластический материал и биологически активные вещества.
Простые белки (протеины) построены только из аминокислот. В состав сложных белков, помимо аминокислот, входят нуклеиновая и фосфорная кислоты, углеводы и ряд других веществ. Белки не синтезируются организмом из других макронутриентов (углеводов, жиров) и не запасаются впрок в виде резерва, как, например, жиры, поэтому источником их синтеза являются аминокислоты, поступающие с белками продуктов питания. Аминокислоты — это гетерофункциональные соединения, которых известно более 200, но в состав белков входят только 20 постоянно встречающихся. Из них 8 не синтезируются в организме человека, а синтезируются только растениями, поэтому их необходимо получать с пищей.
К таким незаменимым аминокислотам относятся: валин, лейцин, изолейции, треонин, метионин, лизин, фенилаланин и триптофан. В детском организме не синтезируются также гистидин и аргинин. Дефицит всех этих аминокислот не компенсируется за счет активизации биохимических реакций синтеза в организме и приводит к болезненным последствиям.
Биологическая ценность белков, таким образом, определяется количеством и соотношением входящих в них аминокислот. Белки различаются также по усвояемости. Лучше всего (на 96…98 %) усваиваются белки молока и яиц, а белки злаковых и бобовых усваиваются на 70…80 %.
Сравнительный аминокислотный состав некоторых видов зернобобовых и масличных культур представлен в табл. 3.
Белки злаковых культур неполноценны по ряду незаменимых аминокислот, в первую очередь по лизину, метионину, триптофану и треонину. В то же время горох и соя, да и все бобовые, содержат высокий процент лизина.
По своему аминокислотному составу белок сои наиболее приближен к животным белкам, чем и определяется его пищевая ценность.
Под действием физических (в первую очередь температуры), химических и биологических факторов белки денатурируют — меняется их структура, что сказывается на их качестве.
Особое внимание стоит обратить на белок зародыша сои. Он имеет повышенную биологическую ценность, поскольку является концентратом структурных и ферментативных белков, близких по своим свойствам к физиологическим белкам животной ткани. Его усвояемость составляет 91,6 %.
Таблица 3. Содержание незаменимых аминокислот в белках зерновых, бобовых и масличных культур (%)
| Аминокислоты | Пшеница | Рожь | Тритикале | Ячмень | Овес без пленок | Рис | Просо | Кукуруза | Сорго | Горох | Соя | Амарант | Потребность человека* | Молоко коровье | Говядина I категории |
| Лизин | 3,0 | 4,1 | 3,5 | 3,2 | 4,4 | 3,5 | 2,2 | 2,5 | 2,5 | 6,5 | 6,6 | 8,0 | 4,2 | 6,6 | 8,9 |
| Метионин | 1,7 | 1,8 | 1,75 | 1,7 | 2,3 | 2,9 | 2,4 | 2,1 | 1,6 | 1,4 | 1,4 | 2,3 | 2,2 | 2,4 | 3,8 |
| Триптофан | 1,07 | 1,07 | 1,0 | 1,2 | 1,9 | 1,3 | 1,4 | 0,6 | 0,9 | 0,8 | 1,3 | 1,4 | 1,4 | 1,4 | 1,1 |
| Валин | 4,1 | 4,6 | 4,2 | 5,4 | 5,7 | 6,5 | 4,8 | 4,4 | 5,2 | 4,5 | 5,4 | 2,5 | 4,2 | 6,9 | 5,6 |
| Изолейцин | 3,53 | 3,57 | 3,67 | 3,5 | 4,0 | 4,6 | 3,9 | 2,7 | 5,6 | 5,0 | 5,3 | 3,5 | 4,2 | 6,6 | 4,2 |
| Лейцин | 6,38 | 6,36 | 6,88 | 7,2 | 7,7 | 8,0 | 9,6 | 11,2 | 12,7 | 6,5 | 7,9 | 5,7 | 4,8 | 9,9 | 8,0 |
| Треонин | 2,68 | 3,06 | 2,80 | 2,9 | 3,0 | 3,5 | 3,3 | 3,2 | 2,7 | 3,8 | 3,8 | 3,5 | 2,8 | 4,6 | 4,3 |
| Фенилаланин | 4,48 | 4,62 | 4,79 | 5,1 | 5,7 | 5,2 | 4,8 | 4,1 | 4,3 | 4,8 | 5,1 | 5,2 | 2,8 | 4,9 | 7,8 |
По данным ФАО.
Кроме белка, в зародыше сои также имеются различные формы небелкового азота (10…15 % от общего азота) — это такие биологически активные вещества, как аспарагин (0,3…0,6 %), аллантоин (0,7 %), бетаин (0,3…0,6 %), холин (2,6…3,3 %), лецитин (1,25 %), глютамин (0,35…0,5 %).
Такие бобовые культуры, как горох, соя, фасоль, вика, чечевица, кормовые бобы, люпин и другие содержат белка в 2…3 раза больше, чем зерно злаковых. В семенах сои и люпина белки могут достигать 40 %, а в некоторых видах люпина их было обнаружено 61,27 % к весу сухой массы.
Белки бобовых культур хорошо растворимы (в воде и 10 %-ном растворе хлорида натрия), что улучшает их использование организмом животных.
Биологическая ценность для большинства бобовых равна 75…85 %. Некоторые исследователи утверждают, что белки сои равноценны белку молока. Характерной особенностью белка зерна бобовых является аминокислотный состав, почти соответствующий аминокислотному составу кормов животного происхождения. Поэтому зерно бобовых является хорошим белковым дополнителем рационов, содержащих большое количество легкодоступных белков с высокой концентрацией незаменимых аминокислот.
По сравнению со злаковыми в зерне бобовых культур меньше крахмала и больше сахаров. В них мало (за исключением люпина, сои и нута) жира, представленного насыщенными жирными кислотами с низким йодным числом.
В отличие от семян зерновых культур, содержащих преимущественно углеводы, масличные семена очень богаты белками. Наиболее богаты белками (в %): семена подсолнечника — от 26,0 до 29,2; хлопчатника — от 29,5 до 43,5; льна — до 33,8; горчицы — до 30,0; преобладают белки типа глобулинов, альбуминов и меньше глютелинов и нерастворимых.
Значительно меньше в масличных семенах содержится углеводов; так, в ядре семян подсолнечника их будет 8,54 %, хлопчатника — до 14,08 %. Большое количество фосфатидов содержат (в %): соя — до 2, подсолнечник — 0,94, хлопчатник — 0,94, конопля — до 0,88.
Липиды зерна объединяют большое количество разнообразных по составу веществ, основную долю из которых (63…65 %) составляют жиры — простые липиды, причем 70…85 % жиров представлены триацилглицеролами ненасыщенных жирных кислот: олеиновой, линолевой и линоленовой. В силу этого многие растительные масла жидкие.
Кроме того, различают большую группу сложных липидов (фосфатидов), отличающихся от жиров содержанием фосфорной кислоты, и циклических липидов (стеролов и стеридов). Жир содержится главным образом в алейроновом слое и зародыше. Содержание жира в зародыше зерна (в %, в среднем): пшеницы — 14, проса — 22, гречихи — 17, кукурузы — 30. Некоторые из жиров имеют биологически активные вещества — например, сложные липиды: фитин — источник витамина инозита, липид эргостерол — является провитамином D.
В зерне и семенах преобладают лецитин, кефалин. Это фосфатиды, которые вместе с белками образуют липопротеиды. Они, особенно лецитин, — поверхностно-активные вещества, прекрасные эмульгаторы, широко применяются в производстве шоколада, маргарина и в качестве веществ, предохраняющих жиры от окисления и прогоркания. В зерне пшеницы, ржи, ячменя и риса содержится 0,3…0,6 % фосфатидов, в зародыше пшеницы — 1,6 %, в семенах подсолнечника — 0,7…0,8 %, сои — 1,6…2 %, в зародыше сои — 3,15 %.
Витамины в зерне представлены семью водорастворимыми витаминами: В1 (тиамин), В2 (рибофлавин), В3 (пантотеновая кислота), В6 (пиридоксин), РР (ниацин), Н (биотин), миоинозит в виде фитина, и тремя жирорастворимыми: витамины А, D и Е (табл. 4). Аскорбиновая кислота (витамин С) в зрелых зернах практически не содержится и появляется лишь при прорастании зерна.
Таблица 4. Содержание витаминов в зерновых, бобовых и масличных культурах (мг на 100 г)
| Культура | Витамины | ||||||||
| Водорастворимые | Жирорастворимые | ||||||||
| Тиамин (В1) | Рибофлавин (В2) | Пантотеновая кислота (В3) | Пиридоксин (В6) | Фолиевая кислота (В9) | Ниацин (РР) | Биотин (Н) | β-Каротин | Токоферол (Е) | |
| Пшеница | 0,54 ± 0,11 | 0,17 ± 0,02 | 1,12 ± 0,03 | 0,44 ± 0,09 | 0,04 ± 0,005 | 5,7 ± 1,2 | 0,01 | 0,014 | 6,1 |
| Рожь | 0,44 | 0,16 ± 0,04 | 1,0 | 0,41 | 0,055 | 1,3 | 0,006 | 0,018 | 5,34 |
| Овес | 0,64 ± 0,12 | 0,16 ± 0,04 | 1,0 | 0,28 ± 0,03 | 0,027 | 1,6 ± 0,1 | 0,015 | 0,02 | 2,8 |
| Ячмень | 0,42 ± 0,08 | 0,17 ± 0,04 | 0,7 | 0,47 ± 0,02 | 0,05 | 7,4 ± 3,2 | 0,009 ± 0,003 | 0,01 | 2,7 |
| Сорго | 0,46 | 0,16 | 1,0 | 0,4 | 0,225 | 3,3 | 0,017 ± 0,007 | — | 2,7 |
| Горох | 0,59 ± 0,23 | 0,15 | 2,1 ± 0,1 | 0,27 | 0,05 | 2,2 | 0,019 ± 0,005 | 0,01 | 9,1 |
| Соя | 0,94 | 0,22 | 1,8 | 0,85 | 0,2 | 2,2 | 0,06 | 0,07 | 17,3 |
| Фасоль | 0,4 | 1,0 | 1,94 | 0,92 | — | 4,0 | — | — | — |
| Лен | 0,88 | 0,23 | 5,4 | 1,25 | — | — | — | 0,01 | — |
| Просо | 0,55 ± 0,2 | 0,38 | — | 0,43 | 0,032 | 2,85 | — | 0,01 | 2,3 |
| Рекомендуемая суточная норма потребления*, мг | 1,2…2,1 | 1,3…2,4 | 6 | 1,8…2,3 | 0,2…0,4 | 14…28 | 0,03… 0,15 | 0,8…1,4 | 8…12 |
| Устойчивость к температурам | Н/у | У | О/у | У | У | У | У | О/у | О/у |
Примечания: * — для взрослого населения; Н/у — неустойчив; У — устойчив; О/у — ограниченно устойчив.
Зерно и зернопродукты считаются основными источниками витаминов РР и Е, группы В. Витамины В1, В2 и В6 содержатся в ржаном и пшеничном хлебе, пшеничном зародыше и отрубях, пантотеновая кислота (В3) — в изделиях из ржи, ниацин (РР) — в пшеничном зародыше и ячменных зернопродуктах, биотин (Н) — в овсе, кукурузе и горохе. К примеру, зародыш пшеницы содержит (в мг на 100 г): В1 — 0,24…6,0, В2 — 0,3…1,45, В3 — 0,3…2,6, В6 — 0,7…30, витамин РР — 3,4…7,5, фолиевую кислоту — 0,3…0,7 и токоферол — 15…30.
Минеральные вещества составляют 2…5% от сухого вещества зерна, основа которого — углерод (45 %), кислород (42 %), водород (6,5 %) и азот (1,5 %), образующие его органическую часть. На количественном и качественном составе минеральных веществ сильно сказываются условия выращивания, в том числе химический состав почв.
В соответствии с количественным содержанием, минеральные вещества в зерне принято делить на следующие:
- макроэлементы — Р, K, Мg, Nа, Са;
- микроэлементы — Fе, Мn, Сu, Zn, Sе, I, Мо, Со и др.;
- ультрамикроэлементы — Сs, Сd, Нg, Аg, Аu, Rа.
Содержание некоторых элементов в зернобобовых и масличных культурах, рекомендуемая среднесуточная норма потребления минеральных веществ для человека приведены в табл. 5.
Таблица 5. Содержание некоторых минеральных элементов в зерне (мг на 100 г)
| Культура | Na | К | Са | Mg | Р | Fe |
| Зерновые | ||||||
| Пшеница твердая | 8 | 325 | 63 | 114 | 368 | 5,3 |
| Рожь | 4 | 424 | 59 | 120 | 366 | 5,4 |
| Гречиха | 4 | 325 | 70 | 258 | 334 | 8,3 |
| Рис | 30 | 314 | 40 | 116 | 328 | 2,1 |
| Сорго | 28 | 246 | 99 | 127 | 298 | 4,4 |
| Кукуруза | 27 | 340 | 34 | 104 | 301 | 3,7 |
| Амарант | 4 | 508 | 159 | 248 | 557 | 7,6 |
| Бобовые | ||||||
| Горох | 33 | 873 | 115 | 107 | 329 | 6,8 |
| Фасоль | 40 | 1100 | 150 | 103 | 480 | 5,9 |
| Соя | 6 | 1607 | 348 | 226 | 603 | 15,0 |
| Нут | 72 | 1084 | 193 | 126 | 444 | 2,6 |
| Масличные | ||||||
| Лен | — | 86 | 500 | 400 | 199 | — |
| Кунжут | 75 | 497 | 1474 | 540 | 720 | 61 |
| Подсолнечник | 160 | 647 | 367 | 317 | 530 | 61 |
| Суточная потребность, мг | 4000…6000 | 2500…5000 | 800…1000 | 300…540 | 1000…1500 | 12…15 |
Зола бобовых на 75 % состоит из фосфора, калия, на долю других элементов приходится 25 % веса золы. По сравнению с зерном злаковых в них меньше магния и больше кальция и особенно серы, поэтому бобовые служат ценным источником этих элементов в кормлении животных. Кроме того, в них имеются марганец, медь, молибден, бор, йод, кобальт, цинк и др.
В зерне бобовых в значительных количествах обнаружены витамины В1 и В2 , РР, А, Е, K, D и С. Витамины группы В находятся преимущественно в оболочках семян, а жирорастворимые (A, D, Е) — в зародышах.
Недостатком некоторых бобовых культур является содержание в них алкалоидов — в семенах люпина (особенно токсичны узколистные и белые сорта), гликозидов — в семенах фасоли и вики некоторых сортов, в сое присутствует несколько подобных веществ.
Рассмотренные показатели характерны для зерна в целом, однако, в силу различий функционального назначения отдельных анатомических частей зерна, их химический состав существенно различается. Основные запасы питательных веществ (белки, жиры и крахмал) сосредоточены в эндосперме и семядолях. Зародыш злаковых и оболочки содержит большую часть витаминов и микроэлементов. Эти же части зерна являются и основным источником пищевых волокон.
Масличные семена богаты различными органическими кислотами, в том числе лимонной, винной, хлорогеновой, щавелевой, малиновой, уксусной, а также различными витаминами и ферментами; так, в семенах подсолнечника содержится провитамин А и ферменты — каталаза, липаза, амилаза; в семенах хлопчатника — витамины Е, группы В и провитамин А. Особенно богаты витаминами и ферментами кукурузные и пшеничные зародыши, а также семена сои. В кукурузных зародышах содержится комплекс витаминов: Е, В1, В2, РР, С, провитамин А, пантотеновая кислота. Все масличные семена содержат красящие вещества — пигменты, придающие маслам различную окраску.
В масличных семенах присутствуют также минеральные вещества, в состав которых входит калий, натрий, кальций, марганец, магний, фосфор, бор и др. Масличные семена в зависимости от ботанического вида и сорта делятся на типы, которые подразделяются (по содержанию примесей и влаги) на классы. Семена горчицы сарептской (сизой) и белой имеют одинаковую шаровидную форму, но различаются размером и цветом. Первые имеют диаметр 1,65 мм, цвет темно-бурокоричневый или желтый, вторые — диаметр 2,26 мм и беловато-желтый цвет. Содержание жира в сарептской горчице выше, чем в белой.
Содержание жира в семенах конопли северных и умеренных районов выше, чем южных. Семена кунжута окрашены во все оттенки от белого до черного. Светлоокрашенные сорта содержат больше жира и расцениваются выше, чем темноокрашенные. Масличный лен включает семена льна-кудряша и межеумка. Семена льна-кудряша имеют наиболее высокое содержание жира, межеумка — несколько меньшее и долгунца — наименьшее.
Просмотров: 262
Высокобелковые продукты — в каких продуктах больше всего белка?
Ключевые высокобелковые продукты — это мясо и рыба. Они содержат до 25 г белка на 100 г. Сравнимое количество имеется и в молочных продуктах. К ним же можно отнести спортивный протеин (продукт переработки молока) — в нем содержится до 70-80 г белка на 100 г.
Кроме этого, белок есть и во многих растительных продуктах — прежде всего, крупах, зерновых и бобовых культурах. Самой высокобелковой считается соя — а текстурированный соевый протеин (его также называют “соевое мясо”) содержит до 50 г белка на 100 г.
Полный список белковых продуктов и таблицы усвоения растительного и животного белка — в материале ниже.
// В каких продуктах много белка?
Большинство видов мяса (начиная от говядины и свинины, заканчивая курятиной) содержат порядка 25-30 г белка на каждые 100 г. Отметим, что подразумевается содержание белка в сыром мясе. Несмотря на то, что протеины не теряются в процессе готовки, при жарке уменьшается вес мяса.
Следом в списке богатой белком еды идут молочные продукты (около 25-30 г на 100 г), затем — орехи (порядка 20 г на 100 г), семена и различные крупы. Самой высокобелковой крупой является гречка — она содержит от 10 до 12 г протеина на 100 г (до варки).
Однако при расчете итогового содержания белка в продуктах важно учитывать и процент его усвоения. Животные протеины усваиваются на 85-99%, а растительные — лишь на 50-60%. Другими словами, цифру содержания белка в крупах на практике можно делить на два.
// Читать дальше:
Содержание белка в мясе
Фактически, жирность мяса связана с содержанием белка — чем меньше в куске мяса жира, тем выше в нем содержание протеина. Другими словами, в 100 г постной говядины будет больше белка, чем в 100 г свиного окорока. В случае с мясом птицы, самым белковым продуктом является куриная грудка — в ней до 30 г белка.
Отдельно упомянем, что мясные полуфабрикаты содержат дополнительные ингредиенты для увеличения объема. Например, в состав котлет входит более жирное мясо и даже хлебные крошки, что снижает итоговое количество белка. Колбаса и сосиски обычно содержат не более 20 г белка на 100 г.
Плюс ко всему, врачи напоминают, что каждые 50 г колбасы и прочих переработанных мясных продуктов, употребляемых ежедневно, на 18% повышают риск развития колоректальных форм рака. Другими словами, диетологи даже не рассматривают вопрос того, чем полезна колбаса — а напоминают о ее вреде. Всемирная Организация Здравоохранения рекомендует полностью исключить из рациона ветчину, колбасы и сосиски — ограничив суммарное употребление красного мяса (в виде вырезки или фарша) до 350 г в неделю.
// колбаса — в чем вред и можно ли ее есть?
Где содержится белок?
Белок — это важнейший компонент метаболизма человека, без которого невозможно поддерживать здоровье и работоспособность. Порядка 30% от суточной калорийности потребляемых продуктов питания должно приходиться именно на белки — или 1.5-2.5 г протеина на кг веса тела.
В свою очередь, белки состоят из различных аминокислот. Всего существует несколько десятков аминокислот, 22 из которых считаются важными, а 8 — незаменимыми для обмена веществ. Четыре наиболее важные аминокислоты входят в состав спортивной добавки BCAA.
// Читать дальше:
Белковые продукты
В различных количествах белки содержатся во всех продуктах питания — их нет только в воде и в масле. Даже ягоды, фрукты и овощи имеют в составе небольшое количество протеина. Однако, как мы упоминали выше, при расчете потребления белка важно учитывать процент его усвоения.
// Таблица содержания белка в продуктах:
| Класс продуктов питания | Содержание белка на 100 г | Примерный процент усвоения |
| Спортивный протеин | 70 — 75 г | 95 – 99% |
| Мясо и рыба | 20 – 30 г | 95 – 99% |
| Молочные продукты | 10 – 30 г | 90 – 99% |
| Яйца | 12 – 15 г | 95 – 99% |
| Орехи | 10 – 25 г | 65 – 70% |
| Крупы и злаковые | 12 – 15 г | 20 – 60% |
| Фрукты и овощи | 2 – 3 г | 65 – 70% |
Молочные продукты
В качестве высокобелковых продуктов питания главную альтернативу мясу и рыбе составляют сыр, творог и прочие молочные продукты. Например, натуральный творог содержит до 15-20 г белка на 100 г — цифра, сравнимая с содержанием протеина в мясе.
Несмотря на то, что большинство видов сыра содержат до 20-30 г белка на 100 г продукта, содержание жира в них также высоко и достигает 20-30% — поэтому сыр необходимо употреблять в умеренном количестве. Замыкает список богатых белком продуктов молоко, содержащее 2-5 г протеина на 100 г, или 7-12 г белка на большой стакан.
| Название продукта | Содержание белка на 100 г | Примерный уровень усвоения белка |
| Сыр твердых сортов | 25 – 30 г | 90 – 95% |
| Сыр мягких сортов | 20 – 25 г | 90 – 95% |
| Сухое молоко | 20 – 25 г | 90 – 95% |
| Сухие сливки | 20 – 25 г | 90 – 95% |
| Творог обезжиренный | 15 – 20 г | 90 – 95% |
| Брынза | 15 – 20 г | 90 – 95% |
| Творог обычный | 10 – 15 г | 90 – 95% |
| Йогурт | 5 – 6 г | 90 – 95% |
| Мороженное | 3 – 5 г | 90 – 95% |
| Молоко | 2 – 5 г | 90 – 95% |
Растительные продукты и крупы
Содержание белка в растительных продуктах зависит от того, какая именно часть растения используется в пищу. Например, плоды и семена (начиная с орехов, заканчивая фасолью), а также зерна (включая крупы и даже муку) содержат больше белка, чем стебли растений (брокколи, салат) и корни (картофель).
Свежие овощи, фрукты и ягоды содержат минимальное количество белка, поскольку основу их массы составляет вода, углеводы и клетчатка. Отдельно отметим, что в картофеле, ровно как и в прочих овощах, содержится не более 2-3 г белка на 100 г — большая тарелка картофельного пюре содержит меньше протеина, чем одно куриное яйцо.
| Название продукта | Содержание белка на 100 г | Примерный уровень усвоения белка |
| Соевый белок | 30 – 50 г | 90 – 95% |
| Грибы сушеные | 20 – 30 г | 70 – 80% |
| Фасоль | 20 – 25 г | 65 – 70% |
| Чечевица | 20 – 25 г | 65 – 70% |
| Сухой горох | 20 – 22 г | 65 – 70% |
| Орехи | 10 – 25 г | 65 – 70% |
| Картофель | 2 – 3 г | 65 – 70% |
| Овощи и фрукты | 2 – 3 г | 65 – 70% |
| Грибы свежие | 1 – 3 г | 65 – 70% |
| Ягоды | 1 – 2 г | 65 – 70% |
Самые белковые крупы
Напомним, что большинство круп относятся к продуктам переработки злаковых культур (прежде всего это пшеница, рис, кукуруза, ячмень и овес). Также из злаковых изготавливается различная мука. Исключениями являются гречка и киноа, являющиеся, по сути, семенами цветков — и именно в таких псевдозлаковых содержится больше белка.
Несмотря на высокое содержание белка в пшенице, примерно третья часть от его общей массы является глютеном — веществом, способным вызывать пищевую аллергию у некоторых людей. Именно содержанием глютена объясняется низкий уровень усвоения белков из пшеничной муки и других продуктов переработки пшеницы.
| Название продукта | Содержание белка на 100 г сухой крупы | Примерный уровень усвоения белка |
| Пшеничная мука | 12 – 15 г | 25 – 30% |
| Макаронные изделия | 12 – 15 г | 25 – 30% |
| Ржаная мука | 10 – 12 г | 30 – 40 % |
| Гречневая крупа | 10 – 12 г | 50 – 60% |
| Овсяная крупа | 10 – 12 г | 50 – 60% |
| Перловая крупа | 10 – 12 г | 50 – 60% |
| Манная крупа | 10 – 11 г | 50 – 60% |
| Кукуруза | 10 – 12 г | 50 – 60% |
| Бурый рис | 2.5 – 3.5 г | 50 – 60% |
| Белый шлифованный рис | 2 – 3 г | 50 – 60% |
// Читать дальше:
Соевый протеин
Соевые бобы — самый белковый растительный продукт. Соя содержит до 50 г белка на 100 г, что вдвое превышает содержание протеина в мясе. Для удобства использования в пищу соевые бобы обычно проходят процедуру переработки, в результате чего изготавливается текстурированная соя, известная как «соевое мясо».
Большинство опасений о вреде сои для здоровья связано с содержанием в ней фитоэстрогены — белки, структурно похожие на эстрогены. Хотя они способны влиять на те же рецепторы организма, уровень этого влияния весьма ограничен.
Исследования не показывают связи между употреблением разумных количеств продуктов из сои на уровень эстрогенов в организме здоровых мужчин. Однако ведутся исследования на тему возможного негативного воздействия фитоэстрогенов на организм подростков.
// Как эстрогены (гормоны женственности) влияют на организм мужчин?
***
Больше всего белка содержится в животных продуктах — мясе, рыбе и молочке. Лидерами по содержанию белка в растительных продуктах является соя, фасоль и чечевица. Содержание белка в большинстве круп варьируется в пределе 10-12 г на 100 г сухой крупы, однако уровень его усвоения составляет 50-60% — против 90-95% усвоения белка из мяса.
Научные источники:
- Protein Intake – How Much Protein Should You Eat Per Day?, source
- Protein Digestibility Corrected Amino Acid Score, source
В продолжение темы
Дата последнего обновления материала — 7 января 2021
Содержание белков в бобовых
Категория продуктов
Все продукты Мясо Мясо убойных животных Мясо диких животных (дичь) Субпродукты Мясо птицы (и субпродукты) Рыба Морепродукты (все категории) Моллюски Ракообразные (раки, крабы, креветки) Морские водоросли Яйца, яичные продукты Молоко и молочные продукты (все категории) Сыры Молоко и кисломолочные продукты Творог Другие продукты из молока Соя и соевые продукты Овощи и овощные продукты Клубнеплоды Корнеплоды Капустные (овощи) Салатные (овощи) Пряные (овощи) Луковичные (овощи) Паслёновые Бахчевые Бобовые Зерновые (овощи) Десертные (овощи) Зелень, травы, листья, салаты Фрукты, ягоды, сухофрукты Грибы Жиры, масла Сало, животный жир Растительные масла Орехи Крупы, злаки Семена Специи, пряности Мука, продукты из муки Мука и отруби, крахмал Хлеб, лепёшки и др. Макароны, лапша (паста) Сладости, кондитерские изделия Фастфуд Напитки, соки (все категории) Фруктовые соки и нектары Алкогольные напитки Напитки (безалкогольные напитки) Пророщенные семена Вегетарианские продукты Веганские продукты (без яиц и молока) Продукты для сыроедения Фрукты и овощи Продукты растительного происхождения Продукты животного происхождения Высокобелковые продукты
Содержание нутриента
ВодаБелкиЖирыУглеводыСахараГлюкозаФруктозаГалактозаСахарозаМальтозаЛактозаКрахмалКлетчаткаЗолаКалорииКальцийЖелезоМагнийФосфорКалийНатрийЦинкМедьМарганецСеленФторВитамин AБета-каротинАльфа-каротинВитамин DВитамин D2Витамин D3Витамин EВитамин KВитамин CВитамин B1Витамин B2Витамин B3Витамин B4Витамин B5Витамин B6Витамин B9Витамин B12ТриптофанТреонинИзолейцинЛейцинЛизинМетионинЦистинФенилаланинТирозинВалинАргининГистидинАланинАспарагиноваяГлутаминоваяГлицинПролинСеринСуммарно все насыщенные жирные кислотыМасляная к-та (бутановая к-та) (4:0)Капроновая кислота (6:0)Каприловая кислота (8:0)Каприновая кислота (10:0)Лауриновая кислота (12:0)Миристиновая кислота (14:0)Пальмитиновая кислота (16:0)Стеариновая кислота (18:0)Арахиновая кислота (20:0)Бегеновая кислота (22:0)Лигноцериновая кислота (24:0)Суммарно все мононенасыщенные жирные кислотыПальмитолеиновая к-та (16:1)Олеиновая кислота (18:1)Гадолиновая кислота (20:1)Эруковая кислота (22:1)Нервоновая кислота (24:1)Суммарно все полиненасыщенные жирные кислотыЛинолевая кислота (18:2)Линоленовая кислота (18:3)Альфа-линоленовая к-та (18:3) (Омега-3)Гамма-линоленовая к-та (18:3) (Омега-6)Эйкозадиеновая кислота (20:2) (Омега-6)Арахидоновая к-та (20:4) (Омега-6)Тимнодоновая к-та (20:5) (Омега-3)Докозапентаеновая к-та (22:5) (Омега-3)Холестерин (холестерол)Фитостерины (фитостеролы)СтигмастеролКампестеролБета-ситостерин (бета-ситостерол)Всего трансжировТрансжиры (моноеновые)Трансжиры (полиеновые)BCAAКреатинАлкогольКофеинТеобромин
Химический состав различных гибридов кукурузы
Кукуруза считается древнейшим культурным растением на земле, не способным к самосеву и одичанию. Будучи важным компонентом питания, уступает кукуруза лишь пшенице и рису.
Биохимические свойства кукурузы делают ее очень полезным продуктом питания. В ее состав входят ценные для человека витамины группы В, РР, С, калий, фосфор, молибден, фтор, йод, медь и другие. В состав зерен кукурузы входят жиры, углеводы, моносахариды, дисахариды, клетчатка, крахмал, зола.
В составе белка кукурузы входят такие незаменимые аминокислоты как лизин и триптофан. А как нам известно, лизин — является одним из лимитирующих кислот, необходимых для усвоения пищевых белков в организме человека [2].
Когда же речь идет о «пищевой» кукурузе, мы подразумеваем только сладкие сорта. На стадии молочной спелости зерна кукурузы обладают мягким и нежным вкусом, это обуславливается пониженным содержанием крахмала и большим содержанием полисахаридов.
Так же в стадии молочной спелости желтозерные сорта кукурузы имеют высокое содержание каротина и минеральных солей, таких как антиоксиданты и каротин. Каротин делает большую работу в нашем организме, например: защищает нас от свободных радикалов, повышает стрессоустойчивость, помогает организму быстрее адаптироваться в непривычных и сложных условиях [4].
Установлено, что кукуруза является легкоусвояемой культурой, имеющей высокую пищевую и биологическую ценность. При включении в рацион кукурузы наблюдается активизация обменных процессов, что благотворно влияет на самочувствие, жизненный тонус, общее оздоровление.
Зерно кукурузы является источником белка для организма человека и животного. В состав зерна кукурузы содержание белка примерно равно 11,8 %. Известно, что белки образуют вязкие коллоидные растворы, что играет не маловажную роль в технологии производства. Например: из нее выходит прекрасный по своим вкусовым свойствам хлеб, блины, лепешки, попкорн, хлопья. Зерна кукурузы подвергают консервированию и используют в ряде блюд. В процессе консервирования кукуруза сохраняет почти все свои полезные вещества. Также кукурузу используют для приготовления крахмала, пищевого масла, комбикормов для сельскохозяйственных животных [1].
Однако, разные гибриды кукурузы обладают неодинаковыми питательными достоинствами, поэтому оценка особенностей гибридов является важной с точки зрения использования кукурузы в комбикормах, хлебе, консервах и так далее.
В Краснодарском крае широко выращивается эта культура, число гибридов, используемых в производстве очень разнообразно. Для оценки питательных особенностей нами были выбрано 11 гибридов. Нами был изучен химический состав, проведено сравнение витаминного состава, также сделана оценка каротиноидного состава по отношению кукурузному глютену. Данные исследований представлены в таблице1.
Таблица 1
Химический состав различных гибридов кукурузы
Название гибрида | Влажность,% | Протеин | Жир | Клетчатка | Кальций | Фосфор | Зола |
Краснодарский 620 СВ | 8,7 | 8,3 | 4,2 | 1,04 | 1,8 | 0,039 | 1,2 |
Краснодарский 395 УСВВ1 | 9,1 | 9,3 | 4,31 | 1,2 | 2 | 0,041 | 1,3 |
Краснодарский 415 МВ | 7 | 9,19 | 4,17 | 1,5 | 2,6 | 0,035 | 0,8 |
Российский 209 МВ | 8,9 | 8,5 | 3,5 | 1,6 | 1,9 | 0,04 | 1,3 |
Краснодарский 632 МВ | 8,6 | 8,7 | 3,95 | 1,61 | 2,5 | 0,04 | 0,9 |
Краснодарский 403 МВ | 8 | 9,2 | 4,15 | 1,67 | 1,9 | 0,045 | 1,31 |
Краснодарский 290 АМВ | 7,9 | 9,5 | 3,3 | 2,1 | 2,5 | 0,05 | 0,7 |
Краснодарский 410 МВ | 7,6 | 8,8 | 4 | 2,1 | 2,07 | 0,043 | 1 |
Российский 109 МВ | 9,31 | 9,3 | 4,1 | 2,01 | 2,4 | 0,045 | 1,3 |
Краснодарский 385 МВ | 7,5 | 9,03 | 3,7 | 2,54 | 1,9 | 0,051 | 0,8 |
Краснодарский 382 СВ | 9,7 | 9,6 | 3,23 | 3,5 | 1,8 | 0,045 | 1,2 |
Из таблицы видно, что влажность гибридов колеблется в пределах от 7 до 9,7 %. Содержание сырого протеина фактически одинаково, вариации содержания жира незначительны. Гибрид Краснодарский 382 СВ содержит большее количество клетчатки, наименьшее ее содержание у гибрида Краснодарский 620 СВ. Содержание кальция, фосфора и золы отличается незначительно.
Влажность зерна — один из наиболее важных показателей его качества. Вода оказывает сильное влияние на само зерно и микроорганизмы на его поверхности. На влажном зерне быстрее развиваются микробы, увеличивается число клещей, насекомых, происходят другие изменения [3].
Влажность — фактор, показывающий долю питательных веществ зерна и длительность его хранения. Чем выше содержание влаги в зерновой массе, тем меньше она содержит питательных веществ и тем быстрее портится. Чрезмерное количество влаги приводит к активации физиологических, физико-химических процессов. Зерно начинает набухать, прорастать, расщепляются высокомолекулярные биополимеры, активизируются ферменты. Снижается натура, сыпучесть зерна, оно становится уязвимым для механических повреждений. Если влажным зерно остается на длительный срок, его хранение и обработка становятся невозможными. В любом случае, выход зерна и качество продукции при использовании влажного сырья снижаются [5].
Внутри самого зерна вода влияет на физические, химические, биологические свойства зерна, которые определяют его ценность. Выделить химически связанную воду можно, только нарушив структуру белков, жиров, углеводов, в состав которых она входит. Молекулы такой воды уже не обладают свойствами растворителя, поскольку связаны с гидрофильными веществами. Удаление связанной воды приводит к изменению технологических особенностей зерна.
От количества воды, содержащейся в муке, зависит способность муки к хранению. Мука является менее стойким при хранении продуктом, чем зерно. Повышенная влажность приводит к активизации микрофлоры муки и активации содержащихся в муке собственных ферментов. Предельно допустимый уровень влажности (14,5–15 %) позволяет поддерживать в муке условия дефицита влаги, предотвращающие развитие процессов микробиологической и биохимической порчи [1].
В жире зародыша зерна кукурузы — много ненасыщенных жирных кислот (линолевой и олеиновой). При относительно низком содержании протеина, по обменной энергии она превосходит все другие виды зерна. По сравнению с другими зерновыми, в кукурузе мало кальция — 0,05 %, клетчатки — 2,2 %.
В процессе хранения (более шести месяцев), жир зародыша кукурузы подвергается окислению, и разрушает витамины входящие в состав вводимых в комбикорм премиксов, поэтому такую кукурузу используют в количестве не более 40 % [4].
Белковые вещества кукурузного зерна в основном состоят из двух белков — зеина и глютелина. Каждый из них составляет примерно 40 % белкового баланса зерна. Зеин относится к неполноценным белкам, так как он не содержит аминокислоты лизина.
Глютелин — по своему аминокислотному составу относится к полноценным белкам. Говоря о белках кукурузы, надо также отметить еще одну их физическую особенность: они не набухают в воде. Это, как мы увидим ниже, очень важное обстоятельство с точки зрения оценки технологических свойств кукурузной муки [2].
Область применения кукурузы:
Народная медицина. В этой области огромная польза кукурузы. Для лечения обычно используют все части растения. Кукурузные рыльца — хорошее мочегонное средство. Кукурузное масло способствует снижению холестерина в крови. Блюда из кукурузной муки рекомендовали употреблять женщинам с диагнозом бесплодие, а также мужчинам для улучшения потенции.
Косметология. Кукуруза используется в косметических средствах, т. к. богата витаминами питающими волосы и восстанавливающими эластичность кожи.
Строительство. Как ни странно, но используют кукурузу и в области строительства, в производстве картона и бумаги, т. к. она является и кормовой культурой. Также из неё изготовляют клей и клейстер.
Сельское хозяйство. Используется в виде силоса для подкормки крупного рогатого скота [3].
Литература:
- Биохимические особенности консервирования перца сладкого и его отходов. Николаенко С. Н., Чистоусова М. С. Научные труды SWorld. 2007. Т. 15. № 3. С. 67–68.
- Определение степени антагонизма между lactobacillus casei и streptococcus sp. Николаенко С. Н., Пахомова Е. Ю., Гамзина Т. Ю. Научные труды SWorld. 2008. Т. 23. № 4. С. 33–34.
- Пигментный комплекс плодов тыквы. Николаенко С. Н., Гамзина Т. Ю., Пахомова Е. Ю. Научные труды SWorld. 2009. Т. 27. № 1. С. 7–10.
- Способ изготовления корма для цыплят-бройлеРОВ. Кощаев А. Г., Николаенко С. Н., Шевченко Л. П., Петенко А. И. патент на изобретение RUS 2289270 25.07.2005
- Способ получения кормовой добавки. Кощаев А. Г., Мигина Е. И., Николаенко С. Н. патент на изобретение RUS 2498608 10.01.2012.
Основные термины (генерируются автоматически): кукуруза, белок, зерно кукурузы, кукурузная мука, организм человека, связанная вода, химический состав.
Зерновые белки — обзор
Белки пшеницы и других зерновых
Традиционно зерновые белки классифицируются в соответствии с фракционированием на основе растворимости в альбуминах (белки, растворимые в воде), глобулинах (белки, растворимые в разбавленных солевых растворах), проламинах. (запасные белки, растворимые в водном спирте) и глютелины (запасные белки, растворимые в разбавленной кислоте или щелочи) (Osborne, 1924). Однако фракционирование по Осборну не обеспечивает четкого разделения белков, различающихся биохимически / генетически или по функциональности (Veraverbeke and Delcour, 2002).
Сегодня, особенно в случае пшеницы, белки преимущественно классифицируются с функциональной точки зрения на неглютеновые и глютеновые белки. Первые белки в основном содержатся во фракциях альбумина и глобулина Осборна, тогда как последние в значительной степени нерастворимы в воде и разбавленных солевых растворах. Белки глютена пшеницы состоят из глиадинов и функционально различных глютенинов. Глиадины образуют очень гетерогенную группу мономерных белков глютена, растворимых в водном спирте и, следовательно, являются основными компонентами фракции проламина Осборна пшеницы.Их молекулярная масса колеблется от 30 до 80 кДа. Дисульфидные связи в глиадинах, если они присутствуют, являются внутримолекулярными (Veraverbeke and Delcour, 2002). Глютенины состоят из гетерогенной смеси полимеров с широким диапазоном молекулярных масс от примерно 80 кДа до миллионов. Большая часть растворима в условиях разбавленной кислоты и составляет фракцию глютелина Осборна пшеницы. Полимеры глютенина состоят из множества субъединиц глютенина (GS), которые поперечно сшиты дисульфидными связями.Следовательно, сульфгидрильные группы GS вовлечены во внутри- или межмолекулярные связи. GS могут быть получены при восстановлении дисульфидных связей и биохимически связаны с глиадинами (Veraverbeke and Delcour, 2002; Goesaert et al., 2005).
Свойства белков глютена позволяют превращать пшеничную муку в вязкоупругое тесто, которое идеально подходит для выпечки хлеба и удерживает углекислый газ, вырабатываемый ферментирующими дрожжами. Эти свойства уникальны, и их нельзя найти даже в злаках, тесно связанных с пшеницей, таких как ячмень и рожь.Благодаря своему большому размеру и образованию непрерывной сетки полимеры глютенина придают тесту прочность (сопротивление деформации) и эластичность, в то время как глиадины, как полагают, действуют как пластификаторы, обеспечивая вязкость / пластичность теста. Кроме того, в структуре теста задействованы как ковалентные, так и нековалентные связи (Bushuk, 1998; Wrigley et al., 1998). Важность дисульфидных поперечных связей хорошо известна, и окислительные процессы очень важны во время образования теста (Wieser, 2003).Во время выпечки белки глютена претерпевают ряд сложных изменений, хотя природа этих изменений плохо изучена. Вероятно, они представляют собой комбинацию изменений гидрофобности поверхности белка, сульфидрил-дисульфидных обменов и образования новых дисульфидных поперечных связей (Jeanjean et al., 1980; Schofield et al., 1983; Weegels et al., 1994; Морель и др., 2002).
(PDF) Белковый профиль зерновых, псевдозерновых и зернобобовых культур
Оптимальное технологическое качество хлебобулочных изделий.Присутствие глютен-активных белков
можно обнаружить во фракции проламина ряда сельскохозяйственных культур (пшеница, ячмень, рожь, овес)
(Palen čárová and G álová 2010). Хишенхубер и др. (2006) утверждают, что Кодекс
Alimentarius ограничивает содержание глютена до 20 мг · кг-1 для естественно безглютеновых продуктов. Для
пищевых продуктов, которые не являются естественно свободными от глютена, установлен максимальный предел содержания глютена
200 мг · кг-1, что соответствует 0.02% глютена (Wieser and Koehler
2008).
Результаты (таблица 2) показывают, что доля альбуминов и глобулинов в зерне составляет
от 21,92% (пшеница) до 88,47% (горох). Содержание альбуминов и глобулинов
в гречке — 53,51%, в амаранте — 52,06%. С другой стороны, самое высокое значение процентного содержания проламинов
было обнаружено в пшенице (33,17%), а самое низкое — в бобовых (1,05%
до 4,49%). Наибольшее содержание глютелина было обнаружено в пшенице (33.17%), а наименьшее — у гороха
(8,95%) и чечевицы (4,49%). Gálo v á et al. (2011a) указаны долевые доли
24,49% для альбуминов и глобулинов, 37,42% для проламинов и 29,46% для глютелинов. В своих исследованиях
Soch a et al. (2010) определили содержание альбуминов и глобулинов
22,62% в пшенице и 56,98% в амаранте. Содержание проламина составляло 3,15% в амаранте
и 33,78% в пшенице, тогда как процентная доля фракции глютелина составляла 34.95%
пшеницы и 23,02% амаранта. Приведенные результаты согласуются с процентным соотношением
отдельных белковых фракций в проанализированных нами образцах. На основании
из вышеизложенного можно сказать, что горох, чечевица и нут могут быть отнесены к безглютеновым культурным растениям и, следовательно,
могут использоваться для целей безглютеновой диеты.
Выводы
Целью данной работы было определить протеиновый профиль анализируемых злаков,
псевдозерновых и бобовых культур и рекомендовать, какие из этих сельскохозяйственных культур подходят для использования
в безглютеновой диете.На основании полученных результатов можно утверждать, что наибольшее содержание сырых белков
было обнаружено в чечевице и нуте (21,5%). Лучшая пищевая ценность
с точки зрения процентного соотношения альбуминов и глобулинов составила
, показанная горохом (88,47%) со значением CNQ 8 571,43%. По соотношению запасных белков
наилучшее технологическое качество показала пшеница с содержанием проламина
, равным 33.17% и содержание глютелина 33,99%. Псевдозерновые и бобовые —
, характеризующиеся тем, что их доминирующей фракцией являются цитоплазматические белки, в то время как они
также имеют чрезвычайно низкое содержание глютен-образующих белков, что позволяет использовать их в
безглютеновой диете для больных. целиакии.
Благодарности
Этот анализ был проведен при поддержке Исследовательского центра AgroBioTech, который был создан
в рамках проекта «Создание Исследовательского центра AgroBioTech», ITMS 26220220180 и проекта KEGA
No.021СПУ-4/2015.
Ссылки
Альварес-Джубете Л., Арендт Е.К., Галлахер Е. 2009: Пищевая ценность псевдозерновых и их растущее использование в качестве
функциональных безглютеновых ингредиентов. Trends Food Sci Tech 21: 106-113
Bojňanská T и Urminská D 2010: Влияние натуральных добавок на белковый комплекс хлеба. Potravinarstvo,
Slovak J Food Sci 4: 1-5
Friedman M 1996: Пищевая ценность белков из различных пищевых источников. Обзор.J Agr Food Chem 44: 6-29
Gajdošová A и Šturdík E 2004: Биологические, химические и питательные характеристики хлебобулочных изделий, связанные со здоровьем.
Nova Biotech Chim 4: 133-154 (на словацком языке)
Gálová Z, Trebichalský A, Palenčárová E 2011a: Протеиновый профиль тритикале (X Triticosecale Witt.), Пшеницы
(Triticum aestivum L.) и ржи. (Secale cereale). Potravinarstvo, словацкий J Food Sci 5: 274-277
Gálová Z, Kečkešová M, Kopálová Z, Chňapek M, Poláčková A 2011b: Качество зерновых, псевдозерновых и
бобовых с точки зрения безглютеновой диеты.Potravinarstvo, словацкий J Food Sci 5: 268-273
Hager A, Wolter A, Jacob F, Zannini E, Arendt E 2012: Пищевые свойства и ультраструктура коммерческих продуктов
без глютена из разных ботанических источников по сравнению пшеничным часам. J Cereal Sci 56: 239-247
52
Аспекты питания зерновых белков и подходы к преодолению их дефицита [и обсуждение] на JSTOR
AbstractНизкая питательная ценность зернового протеина в первую очередь связана с высоким содержанием запасного протеина проламина.Эти белки в целом характеризуются очень высоким содержанием пролина и глутамина и низким содержанием лизина и других незаменимых в питании аминокислот. Злаки различаются по содержанию проламина и лизина. Рис и овес имеют относительно низкое содержание проламина, около 10%, и приемлемое содержание лизина. Пшеница и ячмень содержат 40-45% проламина и около 3,5% лизина, в то время как кукуруза и сорго содержат более 50% проламина в протеине семян, что приводит к содержанию лизина ниже 2.5%. Интенсивный скрининг изменений морфологии эндосперма, белкового состава или содержания лизина привел к обнаружению ряда мутантов с пониженным содержанием проламина и повышенным содержанием лизина и других незаменимых аминокислот. Эти мутанты с высоким содержанием лизина и низким содержанием проламина обладают значительно улучшенной питательной ценностью белка семян. Однако все обнаруженные до сих пор мутанты также характеризуются пониженным содержанием крахмала и урожайностью зерна, а разновидности с высоким содержанием лизина были выращены лишь в очень ограниченной степени.В настоящее время изучаются основные подходы к повышению питательной ценности зерновых белков: (i) замена части проламина богатыми лизином запасными белками, присутствующими в небольших количествах в семенах, и (ii) повышение пищевой ценности проламина. генной инженерией.
Информация для издателяКоролевское общество — это самоуправляемое товарищество многих самых выдающихся ученых мира, представляющих все области науки, техники и медицины, и старейшая научная академия, которая постоянно существует.Основная цель Общества, отраженная в его учредительных документах 1660-х годов, заключается в признании, продвижении и поддержке передового опыта в науке, а также в поощрении развития и использования науки на благо человечества. Общество сыграло роль в некоторых из самых фундаментальных, значительных и изменяющих жизнь открытий в истории науки, и ученые Королевского общества продолжают вносить выдающийся вклад в науку во многих областях исследований.
Запасные белки семян зерновых: структура, свойства и роль в использовании зерна | Журнал экспериментальной ботаники
227″> Введение
Зерновые — самые важные сельскохозяйственные культуры в мире, с общей годовой урожайностью зерна, превышающей 2000 миллионов тонн (т), по сравнению с менее чем 250 т для семян бобовых (включая бобовые, сою и арахис) (ФАО, 1999).Хотя выращивается ряд видов зерновых, на три (кукуруза (604 т в 1998 г.), пшеница (589 т в 1998 г.) и рис (563 т в 1998 г.)) вместе приходится более 70% от общего объема производства. К другим зерновым относятся ячмень, сорго, просо (которое включает ряд мелкосемянных тропических видов), овес и рожь в порядке уменьшения общего производства.
Зерна злаков содержат относительно мало белка по сравнению с семенами бобовых, в среднем около 10–12% сухого веса. Тем не менее, они обеспечивают более 200 мт белка для питания людей и домашнего скота, что примерно в три раза превышает количество, полученное из более богатых белком (20–40%) семян бобовых.Помимо своей питательной ценности, белки семян зерновых также влияют на использование зерна в пищевой промышленности. Это особенно важно для пшеницы, которая в значительной степени потребляется людьми после переработки в хлеб и другие продукты. Поэтому неудивительно, что белки семян зерновых были главной темой исследований в течение многих лет с целью понимания их структуры, контроля синтеза и роли в использовании зерна.
232″> Хранение глобулинов Эмбрион и внешний алейроновый слой эндосперма содержат запасные белки глобулина, а белки из эмбрионов кукурузы были охарактеризованы довольно подробно (Kriz, 1989, 1999; Kriz and Schwartz, 1986; Kriz and Wallace, 1991; Wallace and Kriz, 1991 ). Эти белки легко растворимы в разбавленном солевом растворе и имеют коэффициент седиментации около 7.Они имеют ограниченное сходство последовательностей с 7S вицилинами бобовых и других двудольных растений и могут быть гомологичны им; они также имеют схожие структуры и свойства (Kriz, 1999). Родственные белки были обнаружены в зародышах и / или слоях алейронов пшеницы, ячменя и овса (Burgess and Shewry, 1986; Yupsanis et al. ., 1990; Heck et al. ., 1993). 7S-глобулины из зародышей риса также были охарактеризованы (Horikoshi and Morita, 1975), но их связь с 7S-глобулинами других растений не установлена.Глобулины 7S хранятся в белковых телах и, по-видимому, действуют исключительно как запасные белки. Однако они, по-видимому, не являются абсолютно необходимыми для нормального функционирования семян, по крайней мере, у кукурузы, где нулевой мутант нормально ведет себя с точки зрения развития и прорастания (Kriz and Wallace, 1991). Более того, хотя алейрон и зародыш богаты белками по сравнению с крахмалистым эндоспермом, глобулины в этих тканях имеют ограниченное влияние на свойства зерна конечного использования. В мелкозернистых злаках, таких как пшеница, алейрон и зародыш составляют лишь около 10% от сухой массы зерна и обычно удаляются измельчением (пшеница), полировкой (рис), обработкой жемчуга (ячмень) или декортикацией (сорго) перед потребление человеком.Напротив, зародыш кукурузы составляет 10–11% зерна, и высокое содержание в нем белка и масла важно для питания скота.
Накопительные глобулины 11–12S, расположенные в крахмалистом эндосперме, также присутствуют по крайней мере в некоторых зернах злаков. Фактически, в овсе и рисе эти белки образуют основную фракцию запасного белка эндосперма, составляющую около 70–80% от общего белка. В настоящее время известно, что эти белки относятся к широко распространенным глобулинам «бобового» типа, которые встречаются у большинства двудольных видов (Casey, 1999).Белки риса плохо растворяются в разбавленных солевых растворах и, следовательно, классически определяются как глютелины, но они явно принадлежат к семейству глобулинов 11–12S. Они состоят из субъединиц M r ок. 55 000, которые посттрансляционно расщепляются с образованием кислых ( M r примерно 33 000 в овсе, 28–31 000 в рисе) и основных ( M r примерно 23 000 и 20–22 000 соответственно) полипептидные цепи, связанные одной дисульфидной связью (Shotwell, 1999; Takaiwa et al ., 1999). Глобулин овса также напоминает бобовые в плане образования гексамерной структуры с коэффициентом седиментации около 12.
Белки, относящиеся к бобовым, называемые «тритицинами», присутствуют в крахмалистом эндосперме пшеницы, хотя на их долю приходится только около 5% общий белок семян (Singh et al ., 1988). Тритицины состоят из больших ( M r около 40 000) и малых ( M r около 22–23 000) полипептидных цепей, но субъединицы, по-видимому, образуют димерные структуры, а не типичные гексамеры бобовых (Singh и др. ., 1988, 1993; Сингх и Шеперд, 1985).
Высокое содержание запасных белков глобулина в зерне овса может способствовать высокой питательной ценности по сравнению с другими зерновыми культурами, такими как ячмень и пшеница, что является важным фактором с учетом широкого использования овса в качестве корма для скота (Lockhard and Hurt, 1986; Cuddeford, 1995).
Запасные белки проламина: общие свойства
За исключением овса и риса, основными запасными белками эндосперма всех злаков являются проламины.Это название первоначально было основано на наблюдении, что они обычно богаты пролином и амидным азотом, полученным из глютамина, но теперь известно, что комбинированные пропорции этих аминокислот фактически варьируются от 30 до 70% от общего количества для разных зерновых и белковые группы. Аналогичным образом, хотя изначально проламины определялись как растворимые в смесях спирт / вода (например, 60–70% (об. / Об.) Этанола, 50–55% (об.) Пропан-1-ола или пропан-2-ола), некоторые встречаются в нерастворимых в спирте полимерах. Тем не менее, все индивидуальные полипептиды проламина растворимы в спирте в восстановленном состоянии.Молекулярные массы проламинов сильно различаются — от 10 000 до почти 100 000.
Таким образом, очевидно, что запасные белки проламина гораздо более разнообразны по структуре, чем глобулины 7S и 11 / 12S, и возможно, что основные группы проламинов у Triticeae (пшеница, ячмень, рожь) и Panicoideae (кукуруза, сорго, просо) имеют отдельное эволюционное происхождение. Тем не менее, большинство проламинов имеют две общие структурные особенности. Первый — это наличие отдельных регионов или доменов, которые принимают разные структуры друг для друга и могут иметь разное происхождение.Второй — это наличие аминокислотных последовательностей, состоящих из повторяющихся блоков на основе одного или нескольких коротких пептидных мотивов или обогащенных конкретными аминокислотными остатками, такими как метионин. Эти особенности ответственны за высокую долю глутамина, пролина и других специфических аминокислот (например, гистидина, глицина, метионина, фенилаланина) в некоторых группах проламина.
Суперсемейство проламинов
Обсуждение структуры и свойств проламина может сбить с толку неспециалиста из-за сложности фракций и их специальной номенклатуры.Однако наличие полных аминокислотных последовательностей представителей всех основных групп проламинов позволило переопределить их классификацию в отношении структурных и эволюционных взаимоотношений (Shewry and Tatham, 1990).
Эта новая система классификации относит все проламины Triticeae (пшеница, ячмень и рожь) к трем широким группам: богатые серой (S-богатые), бедные серой (S-бедные) и высокомолекулярные (HMW). ) проламины, с несколькими подгруппами в группе S-богатых (таблица 1).Эти группы не соответствуют непосредственно полимерным и мономерным фракциям в пшенице (глютенины и глиадины соответственно), признанным химиками зерновых, поскольку встречаются как мономерные, так и полимерные формы S-богатых и S-бедных проламинов.
Структуры типичных S-богатых, S-бедных и HMW проламинов пшеницы обобщены на рис. 1. Все они содержат обширные повторяющиеся последовательности, основанные на богатых пролином и богатых глутамином мотивах с повторяющимися мотивами S-богатых и богатых глутамином мотивов. Группы S-бедных явно связаны между собой.Сходным образом сходство последовательностей явно присутствует между неповторяющимися доменами S-богатых и HMW проламинов, особенно в положениях консервативных остатков цистеина и соседних с ними аминокислотных остатков. На основании таких сравнений можно сделать вывод, что проламины S-богатые, S-бедные и HMW имеют общее эволюционное происхождение. Более широкие сравнения показывают дальнейшие эволюционные и структурные отношения с несколькими группами белков зеина (см. Ниже), проламинами овса и риса, запасными белками 2S-альбумина двудольных семян, ингибиторами α-амилазы и трипсина семян злаков и диапазоном низких значений M r богатые цистеином растительные белки, включая белки-переносчики липидов и пуроиндолины зерновых культур.Таким образом, эти белки вместе определены как суперсемейство злаковых проламинов растительных белков (Kreis et al. ., 1985).
В пшенице проламины образуют основные компоненты белковой фракции глютена, которая образует вязкоупругую сеть в тесте и в значительной степени отвечает за способность обрабатывать пшеницу для получения хлеба, макаронных изделий и многих других пищевых продуктов.
Рис.1.
Схематические структуры типичных HMW, S-богатых и S-бедных проламинов (на основе последовательностей в Anderson et al ., 1989; Bartels и др. , 1986; Ся и Андерсон, 2001). Повторяющиеся последовательности заштрихованы, а дисульфидные связи между консервативными остатками цистеина (1-8) в γ-глиадине показаны линиями. SH обозначает положения остатков цистеина в проламинах HMW.
Рис. 1.
Схематические структуры типичных HMW, S-богатых и S-бедных проламинов (на основе последовательностей в Anderson et al. ., 1989; Bartels et al. ., 1986; Hsia and Anderson, 2001 ). Повторяющиеся последовательности заштрихованы, а дисульфидные связи между консервативными остатками цистеина (1-8) в γ-глиадине показаны линиями.SH обозначает положения остатков цистеина в проламинах HMW.
Таблица 1. Сводка типов и характеристик проламинов зерна пшеницы (белков глютена)
Компоненты
M r (% всего)
Полимеры или мономеры
Частичный аминокислотный состав (мол.%)
Компоненты
M r (% всего)
Полимеры или мономеры% состав
Таблица 1. Сводка типов и характеристик проламинов зерна пшеницы (белков глютена)
Компоненты
M r (% всего)
Полимеры или мономеры
Частичный аминокислотный состав (мол.%) )
Компоненты
M r (% всего)
Полимеры или мономеры
Проламины кукурузы
Проламины кукурузы (называемые зеинами) и других злаков, вызывающих паникоид (например,грамм. сорго и многие просо) состоят из одной основной группы белков (α-зеины) и нескольких второстепенных групп (β, γ, δ-зеины) (Coleman and Larkins, 1999; Leite et al., ., 1999) (рис. 2). Сравнение аминокислотных последовательностей показывает, что все β, γ и δ-зеины являются членами суперсемейства проламинов, но только γ-зеины содержат повторяющиеся аминокислотные последовательности (два или восемь тандемных повторов Pro-Pro-Pro-Val-His. -Лея). Как β-зеины, так и δ-зеины богаты метионином, причем эти остатки сгруппированы в области, близкой к концу C в первом.
Напротив, α-зеины, по-видимому, не связаны ни с какими другими проламинами, кроме проламинов α-типа других злаков, вызывающих паникоид. Они состоят из двух основных подклассов, называемых зеинами 19K и 22K, на основе их M r , определенного с помощью SDS-PAGE, хотя их истинные молекулярные массы составляют 23–24 000 и 26 500–27 000 соответственно. Оба подкласса содержат вырожденные повторы примерно из 20 аминокислотных остатков, девять таких блоков присутствуют в зеинах Z19 и десять — в зеинах Z22 (рис.2).
α-зеины содержат только один или два остатка цистеина на молекулу и присутствуют в зерне в виде мономеров или олигомеров, в то время как все β-, γ- и δ-зеины более богаты цистеином и образуют полимеры.
Рис. 2.
(a) Одномерный SDS-PAGE общих зеинов кукурузы. (b) Схематическая структура α-зеинов кукурузы M r 19 000 (Z19) и M r 22 000 (Z22). (Взято из Shewry and Tatham, 1990, с разрешения.)
Рис. 2.
(a) Одномерный SDS-PAGE общих зеинов кукурузы. (b) Схематическая структура α-зеинов кукурузы M r 19 000 (Z19) и M r 22 000 (Z22). (Взято из Shewry and Tatham, 1990, с разрешения.)
Синтез и депонирование запасных белков семян зерновых
Запасные белки семян злаков вырабатываются секреторным путем и откладываются в дискретных белковых телах. Однако происхождение белковых тел и механизмы, которые определяют путь транспортировки и отложения запасных белков, все еще не полностью изучены.Считается, что запасающие глобулины 7S и 11S, которые присутствуют в эмбрионе и слое алейронов и в крахмалистом эндосперме, соответственно, некоторых злаков, действуют по тому же пути, что и гомологичные белки в семенах двудольных. Таким образом, они синтезируются на мембранах грубого эндоплазматического ретикулума (ER), ко-трансляционно транспортируются в просвет и затем проходят через аппарат Гольджи в определенную популяцию вакуолей для хранения белка, которые отличаются от литических вакуолей, которые также присутствуют в развивающихся семенах. .Точные детали этого процесса были рассмотрены в другом месте (Kermode and Bewley, 1999). Точные механизмы сортировки не совсем понятны, но физическая агрегация внутри Гольджи, по-видимому, важна, приводя к образованию электронно-плотных агрегатов, которые образуют содержимое плотных везикул. Запасные белки глобулина не содержат расщепляемых про-доменов, которые обеспечивают нацеливание на вакуол, но нерасщепляемые сегменты в последовательности зрелого белка могут быть важны (Kermode and Bewley, 1999).
Механизмы транспорта и отложения проламина менее изучены, чем механизмы глобулинов, но могут иметь место два пути. В кукурузе, других злаках-паникоидах (сорго, просо) и рисе проламины, по-видимому, накапливаются непосредственно в просвете ER, что приводит к образованию дискретных белковых тел, окруженных мембраной происхождения ER (Coleman and Larkins, 1999; Muench ). et al ., 1999). У риса это приводит к наличию двух популяций белковых тел: PB-I, который имеет ER-происхождение и содержит проламины, и PB-II, имеющий вакуолярное происхождение и содержащий глобулины / глютелины (рис.3а) (Ямагата и Танака, 1986; Кришнан и др. ., 1986). Окита и его сотрудники также представили доказательства того, что проламины и глобулины / глютелины синтезируются в отдельных субдоменах ER, причем мРНК проламинов нацелены на грубый ER, связанный с развитием проламинсодержащих белковых тел, и мРНК глобулина / глутелина в более типичные цистернальные мембраны ER (Li et al ., 1993 a ; Choi et al ., 2000). Более поздняя работа показывает, что локализация мРНК проламина является результатом связывания со специфическим сайтом тубулина и актинового цитоскелета (Muench et al ., 1998).
Овес похож на рис тем, что в крахмалистых клетках эндосперма содержится высокая доля запасных белков глобулинового типа, но в этом случае глобулины и проламины совместно расположены в одних и тех же белковых телах, а проламины представлены в виде более светлых включений (рис. . 3b) (Lending и др. ., 1989). Авторы предположили, что это является результатом слияния двух популяций белковых тел, происхождения ER (содержащих проламины) и вакуолярного происхождения (содержащих глобулины).
Принимая во внимание, что проламины кукурузы, риса и, вероятно, также овса, по-видимому, накапливаются непосредственно в ER с небольшими или отсутствующими доказательствами транспорта в вакуоль, в настоящее время есть неопровержимые доказательства того, что оба пути транспорта белка и образования белковых тел действуют в пшенице ячмень и, вероятно, также рожь. Доказательства этого были рассмотрены в другом месте (Galili, 1997), но включают мечение проламинов в составе комплексов Гольджи иммунным золотом (см. Также Shewry, 1999), наблюдение небольших белковых тел внутри или связанных с ER, ассоциацию ферментов-маркеров ER с белком. тела, полученные субклеточным фракционированием и экспрессией белков дикого типа и мутантных белков в гетерологичных системах.Вывод состоит в том, что некоторые проламины, в основном глиадины, транспортируются через Гольджи в вакуоль хранения белка, тогда как другие, в основном глютенины, задерживаются в ER. Галили с соавторами также предположили, что белковые тела, полученные из ER, впоследствии абсорбируются вакуолями хранения белка в процессе, аналогичном аутофагии. Белковые тела в развивающихся зернах пшеницы также содержат темные включения запасного белка глобулина тритицина (рис. 3c) (Bechtel et al ., 1991), который предположительно транспортируется через Гольджи к вакуолярным белковым тельцам.
Точный механизм слияния белковых тел в пшенице еще предстоит выяснить. Однако конечным результатом является присутствие в зрелых сухих клетках эндосперма непрерывной матрицы белков, которая окружает гранулы крахмала и поглощает остатки других клеточных структур. Эта матрица является основой для образования глютеновой сети, когда пшеничная мука смешивается с водой для образования теста.
Механизмы, которые определяют, удерживаются ли проламины в ЭР или транспортируются через Гольджи в вакуоль, неизвестны, и невозможно распознать ни классические сигналы удержания ЭР (т.е.е. C -концевые тетрапептиды (KDEL или HDEL) или вакуолярные нацеленные последовательности в этих белках. Экспрессия дикого типа и мутантных форм γ-зеина и γ-глиадина в гетерологичных системах продемонстрировала, что богатые пролином повторяющиеся последовательности необходимы для удержания ER (Torrent et al ., 1994; Geli et al ., 1994; Altschuler et al. ., 1993; Altschuler and Galili, 1994), и возможно, что эти области образуют белок-белковые взаимодействия, ведущие к образованию нерастворимых отложений, которые накапливаются непосредственно в ЭПР, а не переносятся в Гольджи. и вакуоль (Coleman, Larkins, 1999; Shewry, 1999).
Окита и его сотрудники предположили существование особого механизма, ведущего к удержанию проламинов в ЭП риса. Это включает взаимодействие с молекулярным шапероном BiP (связывающим белком), который может связывать возникающий полипептид и удерживать его в ER до сборки в белковое тело (Li et al. ., 1993 b ; Muench et al . , 1999). Об этом механизме для других зерновых пока не сообщалось.
Рис.3.
Белковые тельца в развивающихся крахмалистых клетках эндосперма злаков. (а) Рис через 7 дней после цветения, демонстрирующий две популяции белковых телец. PB-I — это сферические везикулы, ограниченные единой мембраной, происходящей из ER, и содержат проламины. PB-II являются аморфными, образуются из вакуолярного отложения и содержат глобулины / глютелины. (Взято из Yamagata and Tanaka, 1986, с разрешения.) (B) Овес через 8 дней после цветения, демонстрирующий окрашенные светом отложения проламина (помеченного коллоидным золотом 10 нм) в виде включений в белковых телах, содержащих глобулины (коллоидное золото 5 нм) .(Взято из Lending et al. ., 1989, с разрешения.) (C) Пшеница через 11 дней после цветения показывает включения тритицина (темное окрашивание, обозначено I) в матрице проламина (M). (Взято из Bechtel и др. ., 1991, с разрешения.) Полоски составляют 1,0 мкМ на (а) и 0,5 мкМ на (б) и (в).
Рис. 3.
Белковые тельца в развивающихся крахмалистых клетках эндосперма злаков. (а) Рис через 7 дней после цветения, демонстрирующий две популяции белковых телец. PB-I — это сферические везикулы, ограниченные единой мембраной, происходящей из ER, и содержат проламины.PB-II являются аморфными, образуются из вакуолярного отложения и содержат глобулины / глютелины. (Взято из Yamagata and Tanaka, 1986, с разрешения.) (B) Овес через 8 дней после цветения, демонстрирующий окрашенные светом отложения проламина (помеченного коллоидным золотом 10 нм) в виде включений в белковых телах, содержащих глобулины (коллоидное золото 5 нм) . (Взято из Lending et al. ., 1989, с разрешения.) (C) Пшеница через 11 дней после цветения показывает включения тритицина (темное окрашивание, обозначено I) в матрице проламина (M).(Взято из Bechtel и др. ., 1991, с разрешения.) Полоски составляют 1,0 мкМ на (а) и 0,5 мкМ на (б) и (в).
Организация белков в белковых телах
Четкое разделение компонентов проламина и глобулина в двухфазных белковых телах овса (рис. 3b) и пшеницы (рис. 3c) может быть результатом начального отложения этих компонентов в отдельных популяциях белковых тел (как в рисе), которые впоследствии предохранитель. Однако это также может быть результатом фазового разделения белков проламина и глобулина из-за их различных структур и свойств.
Доказательства пространственного разделения разных типов проламинов внутри белковых тел менее очевидны, поскольку разные группы проламинов имеют тенденцию проявлять сходные свойства окрашивания при подготовке к электронной микроскопии. Однако Лендинг и Ларкинс представили элегантные исследования, показывающие различия в распределении зеинов внутри белковых тел, связанных с их положением в развивающемся эндосперме (Lending and Larkins, 1989). Таким образом, белковые тела самых молодых клеток в субалейроновой области содержат в основном β- и γ-зеины, которые распределены по всему белковому телу.Проходя в эндосперм, белковые тела увеличивались в размерах с появлением центрально расположенных «локул», содержащих α-зеины. Наконец, эти локулы сливаются, образуя непрерывную центральную область α-зеинов, причем β- и γ-зеины располагаются периферически в зрелых белковых телах центральных клеток эндосперма (Рис. 4).
Анализу развития белковых тел кукурузы способствовала доступность высокоспецифичных антител к α, β и γ-зеинам, и аналогичные исследования для других злаков пока не проводились.Однако недавние исследования, проведенные с различными фракциями глиадина, показывают, что может происходить разделение, приводящее к образованию микрофаз. Очищенные фракции α-глиадина и ω-глиадина растворяли в 70% (об. / Об.) Этаноле, смешивали в соотношениях 1: 3, 1: 1 и 3: 1, наносили на поверхность слюды и давали растворителю испариться. Анализ свойств поверхности высушенных пленок с помощью атомно-силовой микроскопии показал две фазы в пропорциях, приблизительно соответствующих соотношению компонентов (McMaster et al ., 1999) (рис.5). Это говорит о том, что глиадины и глютенины могут разделяться на отдельные микрофазы в белковых телах, что невозможно наблюдать с помощью обычной электронной микроскопии.
Экспрессия в гетерологичных системах также показала, что смесь классов зеина может потребоваться для образования нормальных белковых тел. Таким образом, коэкспрессия γ-зеина необходима для накопления α-зеина в трансгенном табаке (Coleman et al ., 1996), в то время как β- и δ-зеины образуют аномальные белковые тела, когда экспрессируются отдельно в табаке, но, по-видимому, нормальные белковые тела при совместной экспрессии (Bagga et al ., 1995, 1997). Из этих исследований ясно, что еще многое предстоит узнать о механизмах образования белковых тел в зерновых, а также о роли и организации различных белковых групп.
Рис. 4.
Характер развития образования белковых телец в эндосперме кукурузы. Наиболее незрелое белковое тело находится слева, а развитие идет слева направо. Обозначения греческими буквами на нижнем рисунке указывают расположение соответствующих классов зеина, определяемое иммунолокализацией.(Взято из Coleman et al., ., 1999, с разрешения.)
Рис. 4.
Модель развития белковых тел в эндосперме кукурузы. Наиболее незрелое белковое тело находится слева, а развитие идет слева направо. Обозначения греческими буквами на нижнем рисунке указывают расположение соответствующих классов зеина, определяемое иммунолокализацией. (Взято из Coleman et al ., 1999, с разрешения.)
Рис.5.
Топографическое изображение, полученное с помощью сканирующей зондовой микроскопии смеси 1: 1 очищенных α- и ω-глиадинов, нанесенных из раствора на поверхность слюды. Обратите внимание на наличие двух четких фаз. (Взято из McMaster et al., ., 1999, с разрешения.)
Рис. 5.
Топографическое изображение, полученное с помощью сканирующей зондовой микроскопии 1: 1 смеси очищенных α- и ω-глиадинов, нанесенных из раствора на поверхность слюды. Обратите внимание на наличие двух четких фаз. (Взято из McMaster et al ., 1999, с разрешения.)
Пространственное распределение проламинов в крахмалистом эндосперме
Поскольку алейроновые клетки продолжают делиться периклинально в развивающемся эндосперме злаков, самые молодые клетки присутствуют в субалейроновом слое, а самые старые клетки — в центральной части эндосперма. Как у мелкозернистых злаков, так и у видов паникоидов субалейроновые клетки содержат мало гранул крахмала, которые, как правило, меньше, чем в центральных клетках эндосперма.Следовательно, эти клетки содержат большое количество белков, хотя общее содержание белка на клетку мало варьирует по всему эндосперму цельной пшеницы (Evers, 1970). У кукурузы белковые тела в субалейроне и внешних частях эндосперма обогащены γ- и β-зеинами и мало α-зеинов, причем последние более равномерно распределены по эндосперму (Geetha et al ., 1991). Такое распределение согласуется с онтогенезом белковых тел кукурузы, рассмотренным выше.
Различия в распределении белка по эндосперму также встречаются у ячменя (и, вероятно, также у пшеницы), хотя основа для этого неясна.Субалейроновые клетки обоих видов богаты белками, но иммуноцитохимические исследования и исследования ячменя показывают, что они содержат в основном S-богатые и бедные S-образные проламины (в основном гордеины B и C), с только проламином HMW (D гордеин). присутствуют в значительных количествах ниже уровня субалейрона (Shewry et al ., 1996; Tecsi et al ., 2000). Это может иметь значение для использования пшеницы и ячменя, поскольку D-гордеины являются основными компонентами гелевой белковой фракции, которые могут ограничивать модификацию ячменя во время соложения (Smith and Lister, 1983), в то время как гомологичные HMW-субъединицы глютенина пшеницы являются основными. компоненты эластомерных полимеров, которые лежат в основе хлебопечения и других пищевых продуктов (Shewry et al ., 1995).
Регуляция экспрессии гена проламина
Гены проламина являются предметом тканеспецифической регуляции и регуляции развития, экспрессируются исключительно в крахмалистом эндосперме во время среднего и позднего развития, а также регуляции питания, чутко реагируя на доступность азота и серы в зерне (Duffus and Cochrane, 1992). ; Giese, Hopp, 1984). Этот контроль экспрессии генов осуществляется в первую очередь на уровне транскрипции (Bartels and Thompson, 1986; Sørensen et al ., 1989).
Мало что известно о механизмах, с помощью которых экспрессия генов проламина реагирует на серу. Однако был идентифицирован мотив, участвующий в реакции генов проламинов пшеницы, ячменя и ржи с низким содержанием S и богатым S на азот. Этот мотив (мотив N или азотный элемент) (Hammond-Kosack et al ., 1993; Muller and Knudsen, 1993) присутствует в высококонсервативной последовательности, называемой блоком проламина. Блок проламина (иногда называемый элементом эндосперма) был первой регуляторной последовательностью гена проламина, о которой сообщалось, и был идентифицирован путем сравнения промоторов нескольких генов глиадина и гордеина (Forde et al ., 1985). Это выявило присутствие консервативной последовательности, которая имеет длину примерно 30 п.н., примерно на 300 п.н. выше сайта начала транскрипции (сначала она была названа элементом -300). Консенсусная последовательность для элемента: 5′-TGACATGTAA AGTGAATAAG ATGAGTCATG.
Мотив N находится на 3′-конце бокса и имеет консенсусную последовательность G (A / G) TGAGTCAT в S-богатых генах проламина. Он присутствует в обратной ориентации в проламинном боксе S-бедных генов проламина (Shewry et al ., 1999). Он имеет сходство с сайтом связывания фактора транскрипции GCN4, который является компонентом пути передачи сигналов азота у дрожжей и иногда называется GCN4-подобным мотивом (GLM). Коробка проламина содержит второй высококонсервативный мотив, последовательность TGTAAAGT, который был назван эндоспермом или мотивом E (Hammond-Kosack et al ., 1993).
Было показано, что промоторные области, содержащие проламиновый бокс, функционируют при введении конструкций репортерного гена промотор / хлорамфениколацетилтрансфераза (CAT) в трансгенный табак (Colot et al., 1987; Marris и др. ., 1988). Регуляторная роль самого проламинового бокса была установлена ранее (Müller and Knudsen, 1993; Hammond-Kosack et al ., 1993). Мюллер и Кнудсен использовали гомологичную временную систему экспрессии, включающую бомбардировку культивируемых эндоспермов ячменя частицами конструкциями промотор гордеина С / β-глюкуронидаза (GUS). Эти эксперименты подтвердили, что мотивы E и N являются отдельными элементами, и показали, что мотив N оказывает отрицательное влияние на экспрессию гена при низких уровнях азота и взаимодействует с мотивом E и другими вышестоящими элементами, давая высокую экспрессию, когда уровни азота являются адекватными.
Хаммонд-Косак и др. . использовали in vivo для анализа отпечатков пальцев и задержки геля, чтобы показать, что E-мотивы в проламиновом боксе и выше в промоторе гена низкомолекулярной (LMW) субъединицы пшеницы связывают предполагаемый фактор транскрипции, ESBF-1 (Hammond-Kosack ). et al ., 1993). Второй предполагаемый фактор транскрипции, ESBF-II, связывал мотив N до максимальной экспрессии гена. С тех пор было показано, что третий предполагаемый фактор транскрипции, SPA, распознает мотив N (Albani et al ., 1997).
Вместе результаты этих экспериментов подтверждают, что мотив N является важным компонентом механизма регуляции азота для S-богатых и S-бедных генов проламинов. Его функция требует взаимодействия с мотивом E, два мотива вместе составляют проламинный бокс. Однако проламин-бокс присутствует не во всех генах проламина. Промоторы гена зеина, например, содержат высококонсервативный элемент из 15 п.н., который, как предполагается, действует как тканеспецифический энхансер (Quayle and Feix, 1992).Он содержит последовательность TGTAAAG, которая напоминает мотив E, но мотив N отсутствует (Coleman, Larkins, 1999). Мотив N присутствует в промоторах γ-зеина, но он отделен от мотива E, и его функция, если таковая имеется, не исследована (Coleman and Larkins, 1999).
Бокс проламина в целом также не присутствует в промоторах гена проламина HMW (Shewry et al. ., 1999). Вместо этого промоторы проламина HMW содержат главный регуляторный элемент (идентифицированный Thomas and Flavell, 1990), который расположен в последовательности из 38 п.н. с консенсусом: 5′-GTTTTGCAAA GCTCCAATTG CTCCTTGCTT ATCCAGCT.
Расположение этой последовательности высоко консервативно во всех промоторах проламина HMW, начиная с положения от -185 до -189 (Shewry et al. ., 1999). Элемент содержит последовательность TGCAAAG, которая аналогична последовательности E-мотива TGTAAAG, которая также присутствует в генах зеина, но не содержит ничего, напоминающего N-мотив.
Последовательности, соответствующие частям мотивов N и E, присутствуют в промоторах проламина HMW выше основного энхансера (Lamacchia et al., 2001). Однако делеция этих последовательностей, по-видимому, не влияет на активность промотора, по крайней мере, при управлении экспрессией репортерного гена в трансгенном табаке (Halford et al. ., 1989; Thomas and Flavell, 1990).
Белки семян зерновых и использование зерна
Общее содержание белка в семенах зерновых варьируется от 10 до 15% от сухого веса зерна, при этом около половины общего количества составляют запасные белки. Тем не менее, белки имеют большое влияние на конечные потребительские свойства зерна.Проламины, которые образуют основную фракцию запасного белка во всех основных зерновых культурах, за исключением овса и риса, имеют дефицит незаменимых аминокислот лизина, а также треонина и триптофана (особенно в кукурузе). Это приводит к дефициту этих аминокислот в питательных веществах, когда цельное зерно скармливается животным с однокамерным желудком, таким как свиньи и домашняя птица. Поэтому обычно комбинируют злаки с другими источниками этих аминокислот для корма для животных, например, с семенами бобовых (особенно соевых бобов), жмыхом из масличных семян, рыбной мукой или синтетическими аминокислотами.Комбинация семян зерновых и бобовых особенно предпочтительна, поскольку эти два типа семян по существу дополняют друг друга по своему составу незаменимых аминокислот: злаки, как правило, богаты серосодержащими аминокислотами и бедны лизином, а семена бобовых, наоборот.
Пищевая ценность злаков, как правило, не является важным фактором для рациона питания людей в развитых странах, хотя в некоторых развивающихся странах она все еще важна. Основное внимание уделяется влиянию белков зерна на функциональные свойства при переработке пищевых продуктов, поскольку большая часть всех злаков, за исключением риса, потребляется в обработанных пищевых продуктах.Качество обработки особенно важно для пшеницы, где белки глютена являются основным фактором, определяющим качество конечного использования.
Протеины клейковины и качество зерна пшеницы
Большая часть потребляемой людьми пшеницы перерабатывается из белой муки, которую получают путем измельчения для удаления зародыша (зародыша) и отрубей (околоплодник, семенник, нуцеллярный слой и алейроновый слой). Таким образом, он соответствует крахмалистым клеткам эндосперма и содержит большое количество крахмала и глютена.
Белки глютена образуют непрерывный матрикс в зрелых сухих клетках эндосперма, как обсуждалось выше. Когда мука смешивается с водой для образования теста, белковые матрицы в отдельных ячейках объединяются, образуя непрерывную сеть. Это придает вязкоупругие свойства, которые позволяют тесту расширяться путем ферментации и выпекать его в дрожжевой хлеб или перерабатывать в макароны, лапшу и ряд других продуктов.
Молекулярная основа вязкоупругих свойств пшеничной глютена в течение многих лет привлекала ученых-зерновиков как явление, представляющее фундаментальный интерес, а также в отношении улучшения свойств конечного использования пшеничной муки.Особое значение имеют полимеры глютенина, и хорошо известно, что крепкие (т.е. высоковязко-эластичные) тесто содержат высокие доли высокомолекулярных полимеров глютенина (Field et al. ., 1983 b ). Однако прорыв в понимании происходил около 20 лет назад, когда Пейн и его сотрудники продемонстрировали, что аллельные вариации в составе HMW-проламинов (HMW-субъединиц глютенина) сильно коррелировали с различиями в хлебопекарном качестве европейской хлебной пшеницы (Payne , 1987).Эта ассоциация была подтверждена во многих лабораториях по всему миру и привела к детальным исследованиям структуры и свойств субъединиц HMW.
Сорта гексаплоидной мягкой пшеницы имеют шесть генов субъединиц HMW, по два в локусах Glu-1 на длинных плечах хромосом группы 1 (1A, 1B, 1D). Каждый из этих локусов кодирует одну субъединицу x-типа и одну субъединицу y-типа. Однако различия в экспрессии генов приводят к присутствию только трех, четырех или пяти белков субъединиц HMW, причем субъединицы 1Dx, 1Dy и 1Bx присутствуют во всех сортах, а субъединицы 1Ax и / или 1By — только в некоторых сортах.Хорошее качество хлебопечения особенно связано с присутствием субъединицы 1Ax (по сравнению с молчащим или нулевым аллелем) и пары субъединиц 1D, кодируемой хромосомой 1Dx5 + 1Dy10 (по сравнению с парами аллельных субъединиц 1Dx2 + 1Dy12, 1Dx3 + 1Dy12 и 1Dx4 +. 1Ды12).
Результаты ряда исследований согласуются с гипотезой о том, что субъединицы HMW образуют эластомерную полимерную сеть, которая обеспечивает «основу» для взаимодействий с другими субъединицами глютенина и с глиадинами. Нет никаких сомнений в том, что эта сеть стабилизируется межцепочечными дисульфидными связями (Shewry and Tatham, 1997), но Белтон предположил, что межцепочечные водородные связи, образующиеся, в частности, между остатками глутамина, присутствующими в повторяющихся доменах (рис.1), также важны для придания эластичности (Belton, 1999).
Биологическое значение биофизических свойств глютена неизвестно, поскольку белки функционируют в основном как запасные белки зерна и не имеют известной биологической потребности в проявлении вязкоупругости. Однако вполне вероятно, что молекулярные взаимодействия, которые определяют эти свойства, изначально устанавливаются в развивающемся зерне, поскольку белковые тельца, выделенные на средней стадии развития, содержат полимеры глютенина с дисульфидной связью (Field et al ., 1983 a ), которая может демонстрировать вязкоупругость (неопубликованные наблюдения авторов). Точные роли фермента, протеиндисульфидизомеразы, в катализе образования дисульфидных связей между белками глютена в ER и BiP в установлении других белковых взаимодействий остаются неопределенными (Grimwade et al ., 1996; DuPont et al . , 1998; Галили, 1997; Shewry, 1999).
Управление составом субъединиц ВМ и качеством зерна пшеницы
Ряд факторов повлиял на выбор субъединиц HMW в качестве ранней мишени для генной инженерии с целью улучшения качества зерна.Они демонстрируют четкую связь с качеством зерна, как с количественными эффектами, связанными с экспрессией гена (например, субъединицы 1Ax) (Halford et al ., 1992), так и с качественными эффектами, связанными с аллельными различиями в структуре и свойствах субъединиц (например, субъединицы 1Dx5 + 1Dy10). . Кроме того, доступен ряд генов, кодирующих субъединицы, связанные с хорошим и плохим качеством выпечки хлеба, а трансгенные продукты легко идентифицируются с помощью простого SDS-PAGE общих белков семян.
На данный момент четыре лаборатории сообщили об экспрессии трансгенов субъединиц HMW в мягкой пшенице, в каждом случае с использованием либо собственного промотора гена, либо промотора другого гена субъединицы HMW (Blechl and Anderson, 1996; Altpeter et al ., 1996; Barro et al. ., 1997; Альварес и др. ., 2000). Все сообщили об экспрессии трансгенов на уровнях, равных или превышающих уровни эндогенных генов субъединиц HMW. Экспрессия трансгена также оказалась ограничена крахмалистыми клетками эндосперма, и это было подтверждено анализом линий трансгенной пшеницы, в которых промотор гена субъединицы HMW 1Dx5 использовался для управления репортерным геном Uid A, кодирующим β-глюкуронидазу (Gus) ( Lamacchia и др. ., 2001).
Влияние трансгенов субъединиц HMW на прочность теста было определено на выбранных линиях, выращиваемых в теплице (Barro et al ., 1997; Rooke et al ., 1999) и полевых участках (Popineau et al ). ., 2001) с помощью миксографа, который измеряет энергозатраты при замесе теста. Это показало, что экспрессия трансгена субъединицы HMW 1Ax1 на фоне низкого качества действительно приводила к ожидаемому повышению прочности теста. Однако, когда ген 1Dx5 субъединицы HMW был высоко экспрессирован либо на том же фоне, либо на фоне хорошего качества выпечки хлеба, это приводило к неожиданным эффектам.Мука, измельченная на этих линиях, не смогла образовать нормальное тесто при гидратации и перемешивании, что привело к снижению параметров миксографа, которые обычно связаны с прочностью теста. Реологический анализ глютена, выделенного из этих линий, показал, что связность глютеновой сети значительно увеличилась, примерно в 100 раз и в 10 раз на фоне плохого и хорошего качества, соответственно (Popineau et al ., 2001), в результате свойства, аналогичные свойствам глютена, модифицированного трансглутаминазой (Larre et al ., 2000).
Увеличение степени перекрестного сшивания между белками глютена в трансгенных линиях могло быть результатом присутствия остатка цистеина на N -концевом конце повторяющегося домена субъединицы 1Dx5 (рис. 1), поскольку цистеин остатки не присутствуют в эквивалентных положениях в других субъединицах 1Dx. Однако есть также свидетельства того, что аллельные пары субъединиц HMW, такие как 1Dx5 + 1Dy10, существуют в виде димеров в полимерах глютенина (Shewry and Tatham, 1997).Экспрессия высоких уровней субъединицы 1Dx5 в отсутствие эквивалентных количеств субъединицы 1Dy10 может, следовательно, привести к радикальной реструктуризации полимеров глютенина.
Из предварительных результатов, обсужденных выше, ясно, что можно манипулировать структурой и свойствами пшеничного глютена с помощью генной инженерии, хотя текущих знаний о структуре и функциональности глютена все еще недостаточно для надежного прогнозирования результатов. Хлебная пшеница в настоящее время трансформируется рядом генов дикого типа и мутантных генов, кодирующих субъединицы HMW и другие белки глютена.Это должно позволить определить точную роль конкретных белков и структурных особенностей, а также создать линии с улучшенными свойствами, подходящие для включения в программы селекции растений.
Выводы
Запасные белки зерновых имеют огромное значение для определения качества и свойств конечного использования зерна. Понимание структуры этих белков, их биофизических и функциональных свойств, а также биологических механизмов, определяющих их синтез, транспортировку и отложение в зерне, важно для подкрепления будущих попыток улучшить качество конечного использования зерна с помощью генной инженерии.
IACR получает грантовую поддержку от Совета по исследованиям в области биотехнологии и биологических наук Соединенного Королевства.
Список литературы
Албани Д., Хаммонд-Косак MCU, Смит С., Конлан С., Колот В., Холдсворт М., Беван М. В..
1997
. Активатор транскрипции пшеницы SPA: специфичный для семян белок bZIP, который распознает GCN4-подобный мотив в бифакториальном эндоспермальном боксе генов проламина. Заводская ячейка
9
, 171
–184. Альтпетер Э, Василь V, Шривастава V, Василь IK.
1996
. Интеграция и экспрессия гена высокомолекулярной субъединицы глютенина 1Ax1 в пшенице. Nature Biotechnology
14
, 1151
–1159. Альтшулер Ю., Галили Г.
1994
. Роль консервативных цистеинов глиадина пшеницы в его транспорте и сборке в белковые тела в ооцитах Xenopus . Журнал биологической химии
269
, 6677
–6682. Альтшулер Ю., Харел Р., Галили Г.
1993
. Роль N — и C -концевых областей γ-глиадина пшеницы в его транспорте через эндоплазматический ретикулум ооцитов Xenopus . Заводская ячейка
5
, 443
–450. Альварес М.Л., Гельман С., Халфорд Н.Г., Лустиг С., Реджиардо М.И., Рябушкина Н., Шури П., Стейн Дж., Валлехос Р.Х.
2000
. Подавление HMW-глютенинов в трансгенной пшенице, экспрессирующей дополнительные HMW-субъединицы. Теоретическая и прикладная генетика
100
, 319
–327. Андерсон О.Д., Грин ФК, Йип Р.Э., Халфорд Н.Г., Шури П.Р., Мальпика-Ромеро Дж-М.
1989
. Нуклеотидные последовательности двух генов высокомолекулярного глютенина из D-генома гексаплоидной мягкой пшеницы, Triticum aestivum L.резюме. Шайенн. Nucleic Acids Research
17
, 461
–462. Bagga S, Adams H, Kemp JD, Sengupta ‐ Gopalan C.
1995
. Накопление зеина 15 килодальтон в новых белковых телах трансгенного табака. Физиология растений
107
, 13
–23. Багга С., Адамс Х., Родрикес Ф. Д., Кемп Дж. Д., Сенгупта ‐ Гопалан К.
1997
.Совместная экспрессия генов δ- и β-зеина кукурузы приводит к стабильному накоплению δ-зеина в белковых телах, полученных из ER, образованных β-зеином. Заводская ячейка
9
, 1683
–1696. Барро Ф, Рук Л., Бекеш Ф, Гра П., Татхам А.С., Фидо Р., Лаццери П.А., Шури П.Р., Барсео П.
1997
. Трансформация пшеницы генами высокомолекулярных субъединиц приводит к улучшенным функциональным свойствам. Nature Biotechnology
15
, 1295
–1299. Bartels D, Thompson RD.
1986
. Синтез информационных РНК, кодирующих обильные белки эндосперма во время развития зерна пшеницы. Plant Science
46
, 117
–125. Bartels D, Altosaar I, Harberd NP, Barker RF, Thompson RD.
1986
. Молекулярный анализ семейств генов γ-глиадина в сложном локусе Gli-1 мягкой пшеницы ( T.aestivum L.). Теоретическая и прикладная генетика
72
, 845
–853. Беккари.
1745
. Де Фрументо
. De Bononiensi Scientiarum et Artium Instituto atque Academia Commentarii, II. Часть I., 122
–127. Bechtel DB, Wilson JD, Shewry PR.
1991
. Иммуноцитохимическая локализация запасного белка тритицина пшеницы в развивающейся ткани эндосперма. Зерновая химия
68
, 573
–577. Belton PS.
1999
. Об эластичности пшеничной клейковины. Journal of Cereal Science
29
, 103
–107. Blechl AE, Андерсон OD.
1996
. Экспрессия нового гена высокомолекулярной субъединицы глютенина в трансгенной пшенице. Nature Biotechnology
14
, 875
–879. Burgess SR, Shewry PR.
1986
. Идентификация гомологичных глобулинов из зародышей пшеницы, ячменя, ржи и овса. Журнал экспериментальной ботаники
37
, 1863
–1871. Кейси Р.
1999
. Распространение и некоторые свойства глобулинов семян. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 159
–169. Choi S ‐ B, Wang C, Muench DG, Ozawa K, Franceschi VR, Wu Y, Okita TW.
2000
. Мессенджер РНК, нацеленная на запасные белки семян риса, на определенные субдомены ER. Природа
407
, 765
–767. Коулман CE, Ларкинс BA.
1999
. Проламины кукурузы. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 109
–139. Коулман CE, Герман Э.М., Такасаки К., Ларкинс Б.А.
1996
. Γ-зеин кукурузы секвестрирует α-зеин и стабилизирует его накопление в белковых телах эндосперма трансгенного табака. Заводская ячейка
8
, 2335
–2345. Колот V, Роберт Л.С., Кавана Т.А., Беван М.В., Томпсон Р.Д.
1987
. Локализация последовательностей в генах белков эндосперма пшеницы, которые обеспечивают тканеспецифическую экспрессию в табаке. Журнал EMBO
6
, 3559
–3564. Каддефорд Д.
1995
. Овес на корм животным. В: Welch RW, ed. Овес: производство и использование
. Лондон: Chapman & Hall, 321
–368. Даффус К.М., Кокрановский член парламента.
1992
. Структура и состав зерна. В Шури PR, изд. Ячмень: генетика, биохимия, молекулярная биология и биотехнология
.Уоллингфорд: CAB International, 291
–317. DuPont FM, Hurkman WJ, Tanaka CK, Chan R.
1998
. BiP, HSP70, NDK и PDI в эндосперме пшеницы. I. Накопление МРНК и белка в процессе развития зерна. Physiologia Plantarum
103
, 70
–79. Эверс нашей эры.
1970
. Развитие эндосперма пшеницы. Анналы ботаники
34
, 547
–555. ФАО.
1999
. Выпуск Ежегодника ФАО
53
. Field JM, Shewry PR, Burgess SR, Forde J, Parmar S, Miflin BJ.
1983
а . Наличие высокомолекулярных агрегатов в белковых телах развивающихся эндоспермов пшеницы и других злаков. Journal of Cereal Science
1
, 33
–41. Филд Дж. М., Шури ПР, Мифлин Б.Дж.
1983
б . Солюбилизация и характеристика белков глютена пшеницы; корреляция между количеством агрегированных белков и качеством выпечки. Журнал продовольственной науки и сельского хозяйства
34
, 370
–377. Forde BG, Heyworth A, Pywell J, Kreis M.
1985
.Нуклеотидная последовательность гена гордеина B1 и идентификация возможных вышестоящих регуляторных элементов в генах запасных белков эндосперма ячменя, пшеницы и кукурузы. Nucleic Acids Research
13
, 7327
–7339. Галили Г.
1997
. Запасные белки проламина пшеницы и ее родственников. В: Ларкинс Б.А., Василь И.К., ред. Клеточная и молекулярная биология развития семян растений
.Нидерланды: Kluwer Academic Publishers, 221
–256. Geetha KB, Lending CR, Lopes MA, Wallace JC, Larkins BA.
1991
. Модификаторы Opaque-2 увеличивают синтез γ-зеина и изменяют его пространственное распределение в эндосперме кукурузы. Заводская ячейка
3
, 1207
–1219. Гели М.И., Торрент М, Людевид Д.
1994
. Два структурных домена опосредуют два последовательных события в нацеливании γ-зеина: удержание белков в эндоплазматическом ретикулуме и образование белковых телец. Заводская ячейка
6
, 1911
–1922. Гизе Х, Хопп Э.
1984
. Влияние азотного питания на количество гордеина, белка Z и матричной РНК β-амилазы в развивающихся эндоспермах ячменя. Carlsberg Research Communications
49
, 365
–383. Grimwade B, Tatham AS, Freedman RB, Shewry PR, Napier JA.
1996
.Сравнение паттернов экспрессии белков глютена пшеницы и белков, участвующих в секреторном пути развития зерновок пшеницы. Молекулярная биология растений
30
, 1067
–1073. Halford NG, Forde J, Shewry PR, Kreis M.
1989
. Функциональный анализ вышележащих участков молчащего и экспрессированного члена семейства генов белков семян пшеницы в трансгенном табаке. Plant Science
62
, 207
–216. Хэлфорд Н.Г., Филд Дж. М., Блэр Х., Урвин П., Мур К., Роберт Л., Томпсон Р., Флэвелл Р. Б., Татхэм А. С., Шури П. Р..
1992
. Анализ субъединиц глютенина HMW, кодируемых хромосомой 1A мягкой пшеницы Triticum aestivum L., указывает на количественное влияние на качество зерна. Теоретическая и прикладная генетика
83
, 373
–378. Хаммонд-Косак MCU, Холдсворт М.Дж., Беван М.В.
1993
. In vivo след гена низкомолекулярного глютенина (LMWG-1D1) в эндосперме пшеницы. Журнал EMBO
12
, 545
–554. Хек ГР, Чемберлен АК, Хо DT ‐ H.
1993
. Ген глобулина 1 эмбриона ячменя, Beg1 : характеристика картирования хромосомы кДНК и регуляция экспрессии. Молекулярная и общая генетика
239
, 209
–218. Хорикоши М., Морита Ю.
1975
. Локализация γ-глобулина в семенах риса и изменения содержания γ-глобулина во время развития и прорастания семян. Сельскохозяйственная и биологическая химия
39
, 2309
–2314. Hsia CC, Андерсон OD.
2001
. Выделение и характеристика генов ω-глиадина пшеницы. Теоретическая и прикладная генетика
103
, 37
–44. Kermode AR, Бьюли JD.
1999
. Синтез, обработка и отложение белков семян: путь синтеза и отложения белка в клетке. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 807
–841. Крайс М, Форд Б.Г., Рахман С., Мифлин Б.Дж., Шури ПР.
1985
. Молекулярная эволюция запасных белков семян ячменя, ржи и пшеницы. Журнал молекулярной биологии
183
, 499
–502. Кришнан HB, Франчески В.Р., Окита TW.
1986
. Иммунохимические исследования роли комплекса Гольджи в формировании белковых тел в семенах риса Planta
169
, 471
–480. Kriz AL.
1989
. Характеристика глобулинов эмбрионов, кодируемых генами кукурузы Glb . Биохимическая генетика
27
, 239
–251. Kriz AL.
1999
. 7S глобулины злаков. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 477
–498. Криз А.Л., Шварц Д.
1986
. Синтез глобулинов в зародышах кукурузы. Физиология растений
82
, 1069
–1075. Криз А.Л., Уоллес Н.Х.
1991
. Характеристика гена кукурузы Globulin ‐ 2 и анализ двух нулевых аллелей. Биохимическая генетика
29
, 241
–254. Lamacchia C, Shewry PR, Di Fonzo N, Forsyth JL, Harris N, Lazzeri PA, Napier JA, Halford NG, Barcelo P.
2001
. Эндосперм-специфическая активность промотора гена запасного белка в семенах трансгенной пшеницы. Журнал экспериментальной ботаники
52
, 243
–250. Ларре С., Денери-Папини С., Попино И., Дешайес Дж., Лефевр Дж.
2000
. Биохимический анализ и реологические свойства глютена, модифицированного трансглютаминазой. Зерновая химия
77
, 121
–127. Leite A, Neto GC, Vettore AL, Yunes JA, Arruda P.
1999
.Проламины сорго, Coix и просо. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 141
–157. Кредитование CR, Ларкинс BA.
1989
. Изменение зеинового состава белковых тел в процессе развития эндосперма кукурузы. Заводская ячейка
1
, 1011
–1023. Lending CR, Chesnut RS, Shaw KL, Larkins BA.
1989
. Иммунолокализация запасных белков авенина и глобулина в развивающемся эндосперме Avena sativa L. Planta
178
, 315
–324. Li X, Franceschi V, Okita TW.
1993
а . Сегрегация мРНК запасных белков на мембранах грубого эндоплазматического ретикулума мембраны эндосперма риса. Ячейка
72
, 869
–879. Li X, Wu Y, Zhang D ‐ Z, Gillikin JW, Boston RS, Franceschi VR, Okita TW.
1993
б . Биогенез тела белка проламина риса: процесс, опосредованный BiP. Наука
262
, 1054
–1056. Lockhart HB, Hurt HD.
1986
. Питание овсом. В: Webster FH, ed. Овес: химия и технология
.Сент-Пол, Миннесота, США: Американская ассоциация химиков злаков, Inc., 297
–308. Маррис К., Галлуа П., Копли Дж., Крейс М.
1988
. 5′-фланкирующая область гена гордеина В ячменя контролирует тканевую и онтогенетическую экспрессию CAT в растениях табака. Молекулярная биология растений
10
, 359
–366. Макмастер Т.Дж., Майлз М.Дж., Ваннербергер Л., Элиассон А.С., Шури П.Р., Татхэм А.С.
1999
. Идентификация микрофаз в пленках смешанных белков α- и γ-глиадина исследована с помощью атомно-силовой микроскопии. Журнал сельскохозяйственной и пищевой химии
47
, 5093
–5099. Muench DG, Wu Y, Coughlan SJ, Okita TW.
1998
. Доказательства связанного с цитоскелетом сайта связывания, участвующего в локализации мРНК проламина в белковых телах в ткани эндосперма риса. Физиология растений
116
, 559
–569. Мюнх Д.Г., Огава М., Окита Т.В.
1999
. Проламины риса. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 93
–108. Мюллер М., Кнудсен С.
1993
. Азотный ответ промотора С-гордеина ячменя контролируется положительной и отрицательной регуляцией GCN4 и бокса эндосперма. Заводской журнал
4
, 343
–355. Payne PI.
1987
. Генетика запасных белков пшеницы и влияние аллельной изменчивости на качество хлеба. Ежегодный обзор физиологии растений
38
, 141
–153. Popineau Y, Deshayes G, Lefebvre J, Fido RJ, Tatham AS, Shewry PR.
2001
.Агрегация проламинов, вязкоупругость глютена и свойства смешивания трансгенных линий пшеницы, экспрессирующих трансгены HMW субъединицы глютенина 1Ax и 1Dx. Журнал сельскохозяйственной и пищевой химии
49
, 395
–401. Quayle T, Feix G.
1992
. Функциональный анализ области -300 генов зеина кукурузы. Молекулярная и общая генетика
231
, 369
–374. Рук Л., Бекеш Ф, Фидо Р., Барро Ф, Грас П., Татхам А.С., Барсело П., Лазцери П., Шури ПР.
1999
. Избыточная экспрессия белка глютена в трансгенной пшенице приводит к получению высокоэластичного теста. Journal of Cereal Science
30
, 115
–120. Shewry PR.
1999
. Синтез, переработка и отложение белков глютена в развивающемся зерне пшеницы. Cereal Foods World
44
, 587
–589. Shewry PR, Tatham AS.
1990
. Запасные белки проламина семян зерновых: структура и эволюция. Биохимический журнал
267
, 1
–12. Shewry PR, Tatham AS.
1997
. Дисульфидные связи в белках глютена пшеницы. Journal of Cereal Science
25
, 207
–227. Shewry PR, Tatham AS, Barro F, Barcelo P, Lazzeri P.
1995
. Биотехнология выпечки хлеба: раскрытие и управление мультибелковым глютеновым комплексом. Био / Технологии
13
, 1185
–1190. Шури П.Р., Бреннан С., Татам А.С., Уорбертон Т., Фидо Р., Смит Д., Григгс Д., Кантрелл И., Харрис Н.
1996
. Развитие, структура и состав зерна ячменя в зависимости от его свойств конечного использования.В: Зерновые 96
. Материалы 46-й Австралийской конференции по химии злаков, Сидней, сентябрь 1996 г., 158
–162. Shewry PR, Tatham AS, Halford NG.
1999
. Проламины Triticeae. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 35
–78. Шотвелл MA.
1999
. Глобулины овса.В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 389
–400. Сингх Н.К., пастырь К.В.
1985
. Структура и генетический контроль нового класса связанных с дисульфидом белков в эндосперме пшеницы. Теоретическая и прикладная генетика
71
, 79
–92. Сингх Н.К., Шеперд К.В., Лэнгридж П., Груэн Л.К., Скерритт Дж. Х., Ригли К.В.
1988
. Идентификация бобовых белков пшеницы. Молекулярная биология растений
11
, 633
–639. Сингх Н.К., Донован Г.Р., Карпентер Х.С., Скерритт Дж. Х., Лэнгридж П.
1993
. Выделение и характеристика кДНК тритицина пшеницы, позволяющая выявить уникальный повторяющийся домен, богатый лизингом. Молекулярная биология растений
22
, 227
–237. Smith DB, Lister PR.
1983
. Гелеобразующие белки в зерне ячменя и их связь с качеством соложения. Journal of Cereal Science
1
, 229
–239. Соренсен М.Б., Кэмерон-Миллс В., Брандт А.
1989
. Транскрипционная и посттранскрипционная регуляция экспрессии генов в развивающемся эндосперме ячменя. Молекулярная и общая генетика
217
, 195
–201. Takaiwa F, Ogawa M, Okita TW.
1999
. Богатые глютелины. В: Shewry PR, Casey R, eds. Белки семян
. Дордрехт: Kluwer Academic Publishers, 401
–425. Tecsi L, Darlington HF, Harris N, Shewry PR.
2000
. Закономерности отложения и распределения белка в развивающемся и зрелом зерне ячменя. В: Генетика ячменя
, VIII. Материалы 8-го Международного симпозиума по генетике ячменя, Аделаида, Австралия, Vol.2, 266
–268. Томас М.С., Флавелл РБ.
1990
. Идентификация энхансерного элемента для специфической для эндосперма экспрессии высокомолекулярного глютенина. Заводская ячейка
2
, 1171
–1180. Torrent M, Geli MI, Ruiz ‐ Avila L, Canals J, Puigdomènech P, Ludevid MD.
1994
. Роль структурных доменов в удерживании γ-зеина кукурузы в ооцитах Xenopus . Планта
192
, 512
–518. Уоллес NH, Криз А.Л.
1991
. Нуклеотидная последовательность клона кДНК, соответствующая гену кукурузы Globulin-2 . Физиология растений
95
, 973
–975. Ямагата Х, Танака К.
1986
. Место синтеза и накопления запасных белков риса. Физиология растений и клеток
27
, 135
–145. Yupsanis T, Burgess SR, Jackson PJ, Shewry PR.
1990
. Характеристика основного белкового компонента алейроновых клеток ячменя Hordeum vulgare L. Journal of Experimental Botany
41
, 385
–392. © Общество экспериментальной биологии
зерна и питание | Совет по питанию зерновых и бобовых
Зерновые и зерновые продукты являются основными продуктами питания культур во всем мире и вносят важный вклад в удовлетворение ежедневных потребностей в питательных веществах с момента начала культивирования около 10 000 лет до нашей эры.Их потребление поощряется диетическими рекомендациями как в Австралии, так и во всем мире из-за значительного вклада, который эти продукты вносят в потребление питательных веществ.
Зерновые злаки с высоким содержанием углеводов, низким содержанием жира, являются хорошими источниками белка и содержат различное количество клетчатки, витаминов и минералов. Питательный состав зерна может варьироваться в зависимости от сорта и условий выращивания в окружающей среде. Содержание белка в пшенице в среднем выше, чем в других злаках.
Зерновые продукты, как цельнозерновые, так и рафинированные, вносят важный вклад в потребление питательных веществ австралийцами. Зерновые продукты являются основными поставщиками семи ключевых питательных веществ в австралийской диете — клетчатки, железа, магния, йода, углеводов и витаминов группы B, включая фолиевую кислоту и тиамин.
В Австралийских диетических рекомендациях перечислены отличительные и важные питательные вещества для зерновых продуктов, включая углеводы, белок, железо, пищевые волокна, йод, магний, цинк, витамины группы B, тиамин, фолат, рибофлавин и ниацин, а также витамин E.Псевдозерновые содержат такое же количество этих питательных веществ, как и зерна, поэтому обычно считаются цельнозерновыми.
Для сравнения, масличные семена, такие как семена чиа , по питательности отличаются от зерновых и не считаются зернами или цельнозерновыми. В них больше масла и меньше углеводов, чем в зернах. Кроме того, семена, включая чиа, содержат очень мало витаминов и минералов группы B, которые считаются важными питательными веществами в зернах.
Таблицу, сравнивающую содержание питательных веществ в различных зерновых, можно скачать здесь.
Питательный Зерна (диапазон г / 100 г)
Псевдозерна (диапазон г / 100 г) Семена чиа (масличные) г / 100 г
Белок 7-13 г
13 г
20 г
Всего жиров 1 — 9 г
3–7 г
31 г
Углеводы 50-72г
52 — 62 г
44 г
Источники: база данных FSANZ NUTTAB, Национальная база данных по питательным веществам Министерства сельского хозяйства США для стандартных справочных материалов и FoodWorks.
Для просмотра ссылок щелкните здесь.
Для получения дополнительной информации о зерне и питательных веществах перейдите по этим ссылкам:
По сравнению с хлопьями для завтрака — хлопья от Post, Kellogg’s & General Mills
Итак, что вы ели на завтрак сегодня утром? Скорее всего, это были хлопья. Фактически, хлопья — это даже не просто еда для завтрака. Вы могли съесть его на завтрак, обед, ужин или перекусить между ними. Вы могли бы съесть его в миске с молоком, в миске без молока или просто взять пригоршню прямо из коробки.Зерновые — один из тех продуктов, которые можно употреблять в любое время суток. И это тоже имеет смысл. Это максимально быстро и просто, и так уж получилось, что большинство злаков имеют фантастический вкус.
Конечно, не все хлопья для завтрака одинаковы. Несмотря на то, что они поставляются в коробках одинакового размера, с кусочками одинаковой формы и по аналогичной цене, между большинством злаков есть немало различий. Некоторые утверждают, что в них много цельного зерна и клетчатки, что они являются хорошим источником витаминов и минералов, полезны для сердца и холестерина.Другие, однако, просто хотят, чтобы вы увидели их «дурацких» мультяшных персонажей, «забавные» формы и цвета. Тем не менее, самые большие различия заключаются не в передней части ящиков с хлопьями, а, скорее, в задней части. Здесь, друзья мои, можно найти важные отличия.
Чтобы показать это, я собрал бок о бок более 50 самых популярных сухих завтраков от Post, Kellogg’s, General Mills и других, чтобы увидеть, как все они сочетаются друг с другом с точки зрения их питательной ценности. состав и ингредиенты.Также в конце я выбрал лучшие и худшие злаки.
Итак, если вы ели хлопья только из-за того, насколько они вкусные, вот ваш шанс начать принимать во внимание и питание.
* ПРИМЕЧАНИЕ № 1 * В таблице ниже можно сортировать. При щелчке по категории (калории, жиры, углеводы и т. Д.) Данные в этом столбце будут отсортированы от наименьшего к наибольшему. Повторный щелчок по нему отсортирует его от самого высокого до самого низкого.
* ПРИМЕЧАНИЕ № 2 * Ответы на любые вопросы, которые могут возникнуть по поводу всего, что вы видите в этом сравнении злаков, скорее всего, будут даны внизу этой самой страницы.Обязательно ознакомьтесь с ним, чтобы получить разъяснения по всему, что может показаться неясным.
Злаки для завтрака по сравнению с
Сухие завтраки Размер порции
(чашки) калорий Всего жиров
(г) Насыщенные жиры
(г) Углеводы
(г) Волокно
(г) Сахар
(г) Белок
(г) Содержит трансжиры Содержит HFCS Петли Froot
(Kellogg’s) 1 120 1 0.5 26 1 13 1 ДА ДА All-Bran
(Kellogg’s) 0,5 80 1 0 23 10 6 4 НЕТ ДА Apple Jacks
(Kellogg’s) 1 120 0.5 0 28 1 15 1 НЕТ ДА Кукурузные хлопья
(Kellogg’s) 1 100 0 0 24 1 * 2 2 НЕТ ДА Кукурузные хлопья
(Kellogg’s) 1 120 0 0 28 1 * 14 1 ДА НЕТ Crispix
(Kellogg’s) 1 110 0 0 25 1 * 3 2 НЕТ НЕТ Хрустящие отруби с изюмом
(Kellogg’s) 1 190 1 0 45 4 20 3 НЕТ ДА Яичный кленовый сироп из злаков
(Kellogg’s) 1 120 1.5 0,5 22 2 13 2 ДА ДА Хлопья матовые
(Kellogg’s) 0,75 110 0 0 27 1 11 1 НЕТ ДА Размер укуса замороженной мини-пшеницы
(Kellogg’s) 24 печенья 200 1 0 48 6 12 6 НЕТ ДА Honey Smacks
(Kellogg’s) 0.75 100 0,5 0 24 1 15 2 ДА НЕТ Smorz
(Kellogg’s) 1 120 2 0,5 25 1 * 13 1 ДА ДА Булочка с корицей Mini-Swirlz
(Kellogg’s) 1 120 2 0 25 1 12 2 ДА НЕТ Продукт 19
(Kellogg’s) 1 100 0 0 25 1 4 2 НЕТ ДА Рис Криспис
(Kellogg’s) 1.25 120 0 0 29 0 3 2 НЕТ ДА Рис Krispies Treats Cereal
(Kellogg’s) 0,75 120 1,5 0 26 0 9 1 ДА ДА Smart Start Healthy Heart
(Kellogg’s) 1.25 230 3 0,5 46 5 17 7 НЕТ ДА Special K
(Kellogg’s) 1 120 0,5 0 22 1 * 4 7 НЕТ ДА Special K Красные ягоды
(Kellogg’s) 1 110 0 0 25 1 10 3 НЕТ ДА Криспи матовый
(Kellogg’s) 0.75 110 0 0 27 0 12 1 НЕТ ДА Wheaties
(General Mills) 0,75 100 0,5 0 22 3 4 3 НЕТ НЕТ Trix
(General Mills) 1 120 1.5 0 28 1 13 1 НЕТ ДА Lucky Charms
(General Mills) 0,75 110 1 0 22 1 11 2 НЕТ НЕТ Fiber One
(General Mills) 0.5 60 1 0 25 14 0 2 НЕТ НЕТ Cheerios
(General Mills) 1 100 2 0 20 3 1 3 НЕТ НЕТ Взрыв йогурта Cheerios Vanilla
(General Mills) 0.75 120 1,5 0,5 24 2 9 2 НЕТ НЕТ Рис Chex
(General Mills) 1 100 0,5 0 23 0 2 2 НЕТ НЕТ Wheat Chex
(General Mills) 0.75 160 1 0 38 5 5 5 НЕТ НЕТ Corn Chex
(General Mills) 1 120 0,5 0 26 1 3 2 НЕТ НЕТ Honey Nut Cheerios
(General Mills) 0.75 110 1,5 0 22 2 9 3 НЕТ НЕТ Multi Grain Cheerios
(General Mills) 1 110 1 0 23 3 6 2 НЕТ НЕТ Всего
(General Mills) 0.75 100 0,5 0 23 3 5 2 НЕТ НЕТ Reese’s Puffs
(General Mills) 0,75 120 3 0,5 22 1 12 2 НЕТ НЕТ Oatmeal Crisp Crunchy Almond
(General Mills) 1 220 5 0.5 46 4 16 6 НЕТ ДА Kix
(General Mills) 1,25 110 1 0 25 3 3 2 НЕТ НЕТ Golden Grahams
(General Mills) 0,75 120 1 0 26 1 11 2 НЕТ НЕТ Франкен Берри
(General Mills) 1 130 1 0 29 1 14 1 НЕТ НЕТ Граф Чокула
(General Mills) 0.75 110 1 0 23 1 12 1 НЕТ НЕТ Cookie Crisp
(General Mills) 0,75 100 1 0 22 1 11 1 НЕТ НЕТ Cocoa Puffs
(General Mills) 0.75 110 1,5 0 23 1 12 1 НЕТ НЕТ Хрустящие тосты с корицей
(General Mills) 0,75 130 3 0,5 25 1 10 1 НЕТ НЕТ Basic 4
(General Mills) 1 200 3 0.5 43 3 13 4 ДА НЕТ Жизнь
(квакер) 0,75 120 1,5 0 25 2 6 3 НЕТ НЕТ ГОЛЕАН Зерновые
(Каши) 1 140 1 0 30 10 6 13 НЕТ НЕТ Виноградные орехи
(Post) 58 граммов 200 1 0 48 7 4 6 НЕТ НЕТ Пшеница дробленая
(Post) 47 граммов 200 1 0 37 6 0 5 НЕТ НЕТ Какао-галька
(Post) 30 граммов 110 1.5 1 26 3 11 1 ДА НЕТ Фруктовая галька
(сообщение) 30 граммов 110 1 1 26 3 11 1 ДА НЕТ Banana Nut Crunch
(Пост) 59 граммов 240 6 0.5 44 4 12 5 НЕТ НЕТ Медовые пучки из овса
(Post) 30 граммов 120 1,5 0 25 2 6 2 НЕТ НЕТ Гребень для меда
(столб) 32 грамма 120 1 0 27 2 10 2 НЕТ НЕТ Oreo O’s
(Сообщение) 27 граммов 110 2 0.5 22 1 13 1 ДА ДА Waffle Crisp
(Пост) 30 граммов 120 2,5 0 25 1 12 2 ДА НЕТ
* Содержание клетчатки указано как «менее 1 грамма».
** Приведенная выше информация о питании НЕ включает молоко.
Лучшие сухие завтраки
Вот злаки, которые считаются лучшими (то есть самыми полезными). При сборе этих круп я руководствовался следующими требованиями. Прежде всего, они должны были абсолютно не содержать трансжиров. Во-вторых, они не должны были содержать насыщенных жиров. В-третьих, они должны были практически не содержать сахара. Затем, после этого, я начал учитывать содержание клетчатки (чем больше, тем лучше), и добавил в этот список злаки, которые содержали хорошее количество клетчатки (но все же соответствовали первым трем рекомендациям).
Я также обратил внимание на содержание белка, а также на то, содержит ли злак какой-либо HFCS (кукурузный сироп с высоким содержанием фруктозы), хотя я не относился к этим двум так же важно, как к вышеприведенным рекомендациям. Последнее, что я принял во внимание, — это ингредиенты и то, из цельного зерна (хорошо) или из рафинированного (не очень) была сделана каша. Итак, вот мой выбор лучших хлопьев для завтрака в произвольном порядке:
- Cheerios (General Mills)
- Измельченная пшеница (Post)
- Fiber One (General Mills)
- Виноградные орехи (столб)
- Wheaties (General Mills)
- Всего (General Mills)
- ГОЛЕАН Каша (Каши)
- All-Bran (Kellogg’s)
- Пшеница Chex (General Mills)
Худшие хлопья для завтрака
Вот злаки, которые считаются худшими (то есть самыми вредными).Во-первых, в этот список автоматически добавлялись любые злаки, содержащие трансжиры, поскольку трансжиры — самый вредный для здоровья ингредиент, который может быть в злаках. Во-вторых, я посмотрел на содержание сахара. Каждый злак, содержащий 10 или более граммов сахара на порцию (который не был получен из настоящих фруктов), также попал в этот список.
Значит ли это, что хлопья с 9 граммами сахара полезны? Конечно нет. Я просто использовал 10 в качестве минимального количества сахара, когда перечислял самые худшие злаки в этом сравнении.Если перечисленные выше злаки являются лучшими, а эти — худшими, то все, что не входит ни в один из списков, занимает промежуточное положение. Вы должны использовать то, что вы узнали здесь, вместе со своим здравым смыслом, когда решаете, следует ли вам есть одну из этих злаков. Итак, двигаемся дальше, в произвольном порядке, вот мой выбор худших хлопьев для завтрака:
- Honey Smacks (Kellogg’s)
- Кукурузные хлопья (Kellogg’s)
- Apple Jacks (Kellogg’s)
- Франкен Берри (General Mills)
- Trix (General Mills)
- Яичный кленовый сироп из злаков (Kellogg’s)
- Basic 4 (General Mills)
- Сморц (Kellogg’s)
- Oreo O’s (Сообщение)
- Froot Loops (Kellogg’s)
- Слойки с какао (General Mills)
- Граф Чокула (General Mills)
- Waffle Crisp (Пост)
- Reese’s Puffs (General Mills)
- Булочка с корицей Mini-Swirlz (Kellogg’s)
- Криспи замороженные (Kellogg’s)
- Какао-галька (столб)
- Фруктовая галька (Пост)
- Cookie Crisp (Дженерал Миллс)
- Замороженные хлопья (Kellogg’s)
- Голден Грэхэмс (Дженерал Миллс)
- талисманы на удачу (General Mills)
- Рис Krispies лечит хлопьями (Kellogg’s)
Часто задаваемые вопросы о сравнении хлопьев для завтрака
На всякий случай, если что-то в приведенном выше сравнении было немного неясным, вот быстрые вопросы и ответы, которые, надеюсь, все прояснят.
Что означает «Содержит трансжиры»?
Во всех сравниваемых хлопьях для завтрака содержание трансжиров указано как «Трансжиры: 0 граммов». Однако в некоторых случаях некоторые из этих злаков на самом деле содержат трансжиры. Позволь мне объяснить…
По закону FDA требует, чтобы пищевые компании указали содержание трансжиров только в том случае, если их пища содержит 0,5 грамма или более в одной порции. Это означает, что если еда содержит ровно 0,49999 граммов трансжиров на порцию, они могут сказать, что их еда содержит 0 граммов трансжиров.Ага, это безумно глупо. Но так оно и есть. Это означает, что если вы съедите несколько порций такой пищи, вы можете съесть несколько граммов трансжиров, даже не подозревая об этом. И даже если вы съедите только 1 порцию, ЛЮБОЙ трансжир все равно будет слишком большим, независимо от его количества.
К счастью, есть способ определить, содержит ли продукт (в данном случае злак) трансжиры, даже если на этикетке написано, что это не так. Просто посмотрите на ингредиенты. Если вы видите термин «гидрогенизированный» или «частично гидрогенизированный», это означает, что в этой пище действительно присутствуют трансжиры.Итак, для этого сравнения хлопьев для завтрака я буквально прочитал список ингредиентов каждой крупы, чтобы увидеть, ДЕЙСТВИТЕЛЬНО ли они содержат какие-либо трансжиры, и удивительное количество из них содержало. Те, у которых действительно было «ДА» в столбце «Содержащиеся трансжиры», а те, которые на законных основаниях НЕ содержали никаких трансжиров вообще, имеют «НЕТ».
Что такое HFCS?
HFCS означает кукурузный сироп с высоким содержанием фруктозы. Если вы не знаете, что это такое, вы можете узнать об этом больше здесь, здесь и здесь.
В чем дело с размерами порций? Почему некоторые в граммах, а не в чашках?
В большинстве сухих завтраков в качестве единицы измерения размера порции использовались чашки. Однако была группа, которой этого не удалось. Итак, я был вынужден указать размер их порции в той единице, в которой они его указали (в большинстве случаев это были граммы).
Почему некоторые из предполагаемых полезных злаков содержат много сахара?
Вот что нужно иметь в виду, глядя на содержание сахара в этом сравнении.Некоторые злаки содержат много сахара, потому что они являются типичными злаками для нездоровой пищи. Другие, однако, содержат много сахара, потому что содержат фрукты. Очевидно, здесь есть небольшая разница, поскольку сахар в Oreo O’s немного отличается от сахара в отрубях с изюмом. Просто нужно иметь в виду.
Безопасны ли злаки на рисовой основе? Я слышал, что они содержат мышьяк? Я также слышал, что белый рис вреден для здоровья. Стоит ли избегать этих злаков?
Я уже затронул тему мышьяка в рисе… не стесняйтесь проверить это. Что касается второй части этого вопроса, вам обязательно стоит ознакомиться с моим сравнением белого и коричневого риса, чтобы понять, какой из них действительно полезен.
Я немного запутался в ваших списках «лучших» и «худших» злаков. Можете ли вы объяснить их немного лучше?
Быстрое уточнение. В этом сравнении было более 50 злаков, и большая часть из них не попала в список лучших или худших. Это означает, что ни один из этих злаков не подходит под определение ОЧЕНЬ лучших или ОЧЕНЬ наихудших.Это не значит, что они здоровы, и это не значит, что они нездоровы. Это просто означает, что они не на вершине ни в одной из категорий.
Так, например, некоторые злаки, не внесенные в список «лучших», все еще в порядке, а некоторые злаки, не внесенные в список «худших», все еще не очень хороши. Поскольку составление списка «посередине» казалось глупой идеей, я приложил все усилия, чтобы предоставить достаточно информации о питании на протяжении всего этого сравнения, чтобы помочь вам самостоятельно принимать обоснованные решения относительно других злаков.
Откуда вы взяли питательную ценность всех этих злаков?
Все данные о питании, использованные в этом сравнении, взяты непосредственно с официальных веб-сайтов каждой марки зерновых. Так что, если что-то покажется неприятным, займитесь этим с ними. Также имейте в виду, что сами бренды зерновых включают небольшую оговорку с информацией о пищевой ценности, которая была примерно такой: «Эта информация о пищевой ценности может измениться. В этом случае проверьте упаковку в магазине, прежде чем покупать ее. ингредиенты и пищевая ценность.»Итак, я полагаю, что это сравнение злаков содержит тот же отказ от ответственности.
Зерновые и цельнозерновые продукты — Better Health Channel
Цельнозерновые злаки и продукты питания могут снизить риск развития некоторых заболеваний, включая ишемическую болезнь сердца, рак толстой кишки, диабет и дивертикулярную болезнь. Общие злаковые продукты включают хлеб, хлопья для завтрака, рис и макаронные изделия. Виды зерна, цельнозерновые и крупяные
К зерновым относятся пшеница, ячмень, овес, рожь, кукуруза, рис, просо и тритикале.Цельнозерновые включают цельнозерновой или цельнозерновой хлеб или хрустящие хлебцы, темный «семявыводящий» хлеб, цельнозерновые хлопья для завтрака, ростки пшеницы, коричневый рис, воздушные цельнозерновые продукты, булгур, киноа, кус-кус, попкорн и овсянку.
Рафинированные злаки включают пирожные, десерты, белый хлеб, макаронные изделия, кексы, сладкое или соленое печенье, очищенные зерновые хлопья для завтрака, белый рис, блины, вафли и пиццу.
Пищевая ценность цельнозерновых злаков
Цельнозерновые злаки состоят из трех основных частей, в том числе:
- отрубей — внешнего слоя зерна, который содержит клетчатку, омега-3 жирные кислоты, витамины и минералы
- эндосперм — основная часть зерно, которое содержит в основном крахмал
- зародыши — наименьшая часть зерна, содержащая витамин Е, фолиевую кислоту, тиамин, фосфор и магний.
Преимущества цельнозерновых злаков
Цельнозерновые злаки являются богатым источником многих основных витаминов, минералов и фитохимических веществ (соединений, содержащихся в растительной пище, которые имеют значительную пользу для здоровья). Типичный продукт из цельнозерновых злаков:
- с низким содержанием насыщенных жиров, но является источником полиненасыщенных жиров, включая омега-3 линоленовую кислоту
- без холестерина
- с высоким содержанием растворимой и нерастворимой клетчатки и устойчивый крахмал
- отличный источник углеводов
- значительный источник белка
- хороший источник витаминов группы B, в том числе фолиевой кислоты
- хороший источник многих минералов, таких как железо, магний, медь, фосфор и цинк
- хороший источник антиоксидантов и фитохимические вещества, которые могут помочь снизить уровень холестерина в крови.
Защитные химические вещества в цельнозерновых продуктах
Цельнозерновые злаки содержат множество различных фитохимических веществ, которые имеют значительную пользу для здоровья). Эти фитохимические вещества включают:
- лигнанов, которые могут снизить риск ишемической болезни сердца и замедлить или обратить вспять рак у животных.
- фитиновая кислота, которая снижает гликемический индекс (ГИ) пищи. Это может быть важно для людей с диабетом и помогает защитить от развития раковых клеток в толстой кишке
- сапонинов, фитостеринов, сквалена, оризанола и токотриенолов, которые, как было обнаружено, снижают уровень холестерина в крови
- фенольные соединения, обладающие антиоксидантным действием .
При очистке цельного зерна
При очистке зерна (например, для производства белой муки) отрубей и зародышевый слой обычно удаляются, остается только эндосперм. Этот процесс может вызвать значительные потери клетчатки, витаминов, минералов, антиоксидантов и фитохимических веществ из зерна.
Некоторое количество клетчатки, витаминов и минералов можно добавить обратно в очищенные зерновые продукты (например, белый хлеб), что компенсирует потери, но невозможно добавить смесь фитохимических веществ, которая теряется при переработке.В Австралии пшеничная мука, используемая для выпечки хлеба, обязательно должна быть обогащена фолиевой кислотой и тиамином, а соль — йодированной.
Рафинированные злаки часто имеют высокий уровень добавления сахара, жира или соли и, как правило, имеют более высокий ГИ, чем их цельнозерновые эквиваленты. Употребление чрезмерного количества рафинированных злаков может вызвать резкое повышение уровня сахара в крови и сильную реакцию со стороны поджелудочной железы, что со временем в сочетании с другим нездоровым диетическим поведением может привести к развитию диабета.
Цельнозерновые помогают защитить от болезней сердца
Употребление в пищу цельнозерновых продуктов из злаков (особенно с клетчаткой из овса или ячменя) связано с защитным действием от сердечных заболеваний у взрослых. Исследования показали, что высокое потребление цельнозерновых продуктов (не менее 2,5 порций в день) связано с более низким риском сердечно-сосудистых событий. Кроме того, исследование женщин в постменопаузе показало, что шесть или более порций цельнозерновых продуктов в неделю защищают от последствий сердечно-сосудистых заболеваний.
Одним из факторов, способствующих развитию сердечных заболеваний, является высокий уровень холестерина в крови. Было обнаружено, что регулярное употребление в пищу цельнозерновых злаков, богатых растворимой клетчаткой, таких как овес (который содержит бета-глюканы) и псиллиум, значительно снижает количество холестерина в кровотоке и может защищать от развития сердечных заболеваний.
Цельнозерновые и диабет 2 типа
Результаты исследований здоровья медсестер I и II показали, что две порции цельнозерновых злаков каждый день могут снизить риск развития диабета 2 типа на 21 процент.В частности, клетчатка из цельнозерновых злаков может защитить от развития этого состояния. Людям с диабетом также может быть полезно употребление цельнозерновых злаков, что связано с улучшением чувствительности к инсулину и лучшим контролем над диабетом.
Цельнозерновые и контроль веса
Люди с избыточным весом или ожирением, как правило, придерживаются высококалорийной диеты. Продукты с высоким содержанием клетчатки, такие как цельнозерновой хлеб и крупы, могут быть эффективной частью любой программы похудания, поскольку они, как правило, имеют более низкую энергетическую плотность.Это означает, что они обеспечивают меньше килоджоулей на грамм пищи. Пища с высоким содержанием клетчатки переваривается дольше и вызывает чувство сытости, что может препятствовать перееданию. Цельнозерновые продукты также содержат мало насыщенных жиров и более здоровые полиненасыщенные жирные кислоты.
Цельнозерновые, злаки и здоровье кишечника
Продукты с высоким содержанием клетчатки, такие как цельнозерновые злаки, увеличивают движение пищи через пищеварительный тракт. Результатом является увеличение объема стула (фекалий), более мягкий и крупный стул и более частое опорожнение кишечника.Это обеспечивает хорошую среду для полезных кишечных бактерий, одновременно снижая уровень деструктивных бактерий и накопление канцерогенных соединений. Пшеничная клетчатка может связывать определенные токсины и выводить их из толстой кишки с фекалиями.
Диета с высоким содержанием клетчатки, особенно с высоким содержанием нерастворимой клетчатки, была связана со снижением риска развития рака толстой кишки и дивертикулярной болезни (состояния, при котором в стенке кишечника образуются «мешочки»).
Цельнозерновые злаки и рак
Ежедневное употребление одной-трех порций цельнозерновых злаков с высоким содержанием пищевых волокон снижает риск колоректального рака у взрослых.Три порции цельнозерновых продуктов в день (около 90 г) могут снизить риск развития колоректального рака на 20%, а дальнейшее снижение риска возможно при более высоком потреблении цельнозерновых продуктов.
Цельнозерновые злаки рекомендуются для здоровья
Цельнозерновые злаки рекомендуются как часть здорового питания. Австралийское руководство по здоровому питанию рекомендует взрослым в возрасте 19–50 лет ежедневно съедать шесть порций зерновых продуктов, большинство из которых должны быть цельнозерновыми. У пожилых людей в возрасте от 51 до 70 лет мужчины и женщины должны потреблять не менее шести порций и четыре порции зерновых продуктов соответственно.
У взрослых в возрасте 70 лет и старше мужчины и женщины должны потреблять не менее 4,5 порций и три порции зерновых продуктов соответственно. Рекомендуемые количества для детей и подростков зависят от их возраста и пола.
Выбор хлеба
Советы при покупке хлеба:
- При покупке зерновых продуктов ищите такие слова, как «цельнозерновой» или «цельнозерновой».
- Зерновой и прожаренный хлеб более питательны и имеют более низкий ГИ, чем более рафинированный хлеб.
- Некоторые «мультизерновые» виды хлеба готовятся из белой муки с добавлением различных цельнозерновых продуктов.
Зерна (диапазон г / 100 г)
Семена чиа (масличные) г / 100 г
7-13 г
13 г
20 г
1 — 9 г
3–7 г
31 г
50-72г
52 — 62 г
44 г
(чашки)
(г)
(г)
(г)
(г)
(г)
(г)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(Kellogg’s)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(General Mills)
(квакер)
(Каши)
(Post)
(Post)
(Post)
(сообщение)
(Пост)
(Post)
(столб)
(Сообщение)
(Пост)
** Приведенная выше информация о питании НЕ включает молоко.